几种麦芽抗酵母菌粗蛋白含量比较及性质分析
2011-11-06姜晓雷李明达赵长新
姜晓雷,韩 丹,李明达,陈 霞,赵 睿,赵长新
(大连工业大学生物与食品工程学院,辽宁大连116034)
几种麦芽抗酵母菌粗蛋白含量比较及性质分析
姜晓雷,韩 丹,李明达,陈 霞,赵 睿,赵长新*
(大连工业大学生物与食品工程学院,辽宁大连116034)
采用硫酸提取及硫酸铵分级沉淀法对八种国内外麦芽的抗酵母蛋白进行了含量分析,得出国产Danpi-2的抗酵母粗蛋白含量最高,达到338!g/g麦芽;同时对抗酵母蛋白的性质进行了分析,得出分子量为10kDa左右的抗酵母粗蛋白能抵抗100℃的高温,把抗酵母粗蛋白加入到发酵液中,酵母数量大量减少,酵母对葡萄糖的利用下降,pH下降缓慢,当粗蛋白含量达到32!g/mL时,酵母生长抑制率达到了50%。
大麦麦芽,抗酵母蛋白,性质分析
1 材料与方法
1.1 材料与仪器
Gardner麦芽、Baudin麦芽、Metcedfe麦芽、Schooner麦芽、Kenpi-8麦芽、Danpi-2麦芽、Kenpi-7麦芽、Ganpi-5麦芽 普兰店中粮麦芽有限公司提供;酵母接种酵母培养基 1%葡萄糖、0.5%蛋白胨、0.5%酵母膏、0.3%大麦汁;考马斯亮蓝R-250/G-250、硫酸铵、浓硫酸、乙腈、NaOH、葡萄糖、酵母膏、蛋白胨等 均为国产分析纯;丙烯酰胺、甘氨酸Amresco;SDS-PAGE低分子量蛋白质标准 TaKala。
ZPS-250H智能恒温恒湿培养箱 黑龙江东拓仪器制造有限公司;WFJ7200可见光分光光度计尤尼柯仪器有限公司;GTL-16A离心机 上海浦东物理光学仪器厂;JM-250型垂直板电泳仪 上海捷迈科贸有限公司。
1.2 实验方法
1.2.1 抗菌蛋白的制备[6-7,9]将麦芽粉碎后,取50目粉末与0.05mol/L硫酸以1∶4混合,冰浴浸提3h,中间每隔15min搅拌一次,提取结束后,在 4℃,8000r/min离心10min,除去沉淀,上清用0.5mol/L氢氧化钠调pH为7.0,将上清加入硫酸铵使饱和度达到30%,放置2h后,在4℃,10000r/min离心10min,除去沉淀,上清液加入硫酸铵至饱和度为80%,在4℃,10000r/min离心10min,除去上清,用少量去离子水复溶沉淀得抗菌蛋白液,将水复溶的抗菌蛋白液加入排阻1000Da的透析袋中,用去离子水透析过夜,至无硫酸根离子(用氯化钡检测),在 4℃,10000r/min离心10min,去除沉淀,上清液冷冻干燥后加入25%乙腈溶液(即乙腈∶水=1∶4)复溶,在4℃放置1h后10000r/min离心10min去除沉淀,保留上清液,保留的上清液用0.45!m的硝酸纤维滤膜过滤,即为抗菌蛋白液。实验组为将抗菌蛋白液加入到酵母培养基中所得吸光值,空白对照为将25%乙腈溶液(即乙腈∶水=1∶4)加入到酵母培养液中所得吸光值。
1.2.2 抗酵母菌活性测定 对酵母抗菌活性的测定:将酵母接种于酵母培养基中,28℃培养12h,使酵母数量达到1.25×107个/mL。按5%接种量接种至新的酵母培养基中,使酵母数量达到6×105个/mL,加入提取的蛋白液1mL,使蛋白达到一定的含量,培养24h,在600nm下测定吸光值。
抑菌率按如下公式计算:抑菌率(%)=(D-S)/ D×100%,式中,D为空白在600nm吸光值,S为实验组样在600nm吸光值。
1.3 抗菌蛋白含量测定
考马斯亮蓝法测蛋白质含量,详见参考文献[10]。
1.4 SDS-PAGE电泳
将麦芽用0.05mol/L硫酸浸提,采用硫酸铵分级沉淀,透析后的上清液用于电泳分析。电泳技术采用不连续电泳方法,分离胶为5%,浓缩胶为12%,操作过程详见参考文献[11]。
1.5 抗酵母粗蛋白耐热性分析
将用0.05mol/L硫酸浸提,采用硫酸铵分级沉淀,透析后的上清液于100℃水浴放置20min,做SDS-PAGE电泳分析,电泳方法同上。
2 结果与分析
2.1 八种麦芽的抗酵母粗蛋白含量分析
由图1显示不同品种麦芽间的抗酵母粗蛋白含量差异较大。国产Danpi-2的抗酵母粗蛋白含量最高,达到338!g/g;其次是加麦Metcedfe、Schooner和国产Ganpi-5,含量为322、310、312!g/g;国产Kenpi -8、Kenpi-7和澳麦Gardner、Baudin含量比较低,分别为234、269、263、271!g/g。以下抗菌活性实验麦芽都以国产Danpi-2为实验原料。

图1 八种麦芽抗酵母粗蛋白含量比较
图2是八种大麦的粗蛋白电泳图,显示不同品种间的抗酵母粗蛋白含量大体相同,蛋白条带位置基本相同,蛋白分子量分别为40、20、10kDa左右。由于用0.05mol/L硫酸作为浸提液,有效地分解了杂蛋白,而保留了一些结构稳定、耐酸分解的蛋白,提高了抗酵母粗蛋白的提取率[10]。

图2 八种麦芽的粗蛋白SDS-PAGE电泳分析
2.2 抗酵母粗蛋白耐热性分析
如图3所示,经100℃煮沸20min后,与图2相比,三条蛋白泳道减少了一条分子量大概为20kDa的泳道,减少的这条蛋白为TLP[7]蛋白,而保留了分子量40kDa的蛋白(泡沫蛋白Z)[7-8],由于一些蛋白受热变性,导致条带变细。分子量10kDa的小分子蛋白为LTP蛋白[8-9],还有一些分子量更小的如硫堇(分子量5kDa左右)、植物防卫素(分子量5kDa)[9],电泳没有显示出来,它们也能抵抗100℃的高温而不变性。

图3 煮沸20min后的麦芽SDS-PAGE电泳图
2.3 抗酵母粗蛋白的抗菌活性分析
2.3.1 抗酵母粗蛋白对酵母生长及葡萄糖代谢的影响 提取国产Danpi-2的抗酵母粗蛋白加入培养基中,如图4所示。当培养基中粗蛋白达到32!g/mL时,酵母生长明显受到抑制,当培养12h后,抑制率达到了50%左右,而对照组在0~8h时处于迅速增长期,加入抗酵母粗蛋白的酵母培养基,酵母在0~18h时处于对数期,增长缓慢。这说明抗酵母粗蛋白的加入,使得细胞受到攻击而发生变化,影响细胞的增殖,延长细胞的对数期且延缓对数期细胞的增殖。

图4 抗酵母粗蛋白对酵母生长的影响
图5显示了酵母生长过程中葡萄糖含量的变化。对照组的糖消耗速率随时间变化迅速下降,到12h时发酵液中的糖已耗完,而添加32!g/mL抗酵母粗蛋白的酵母培养基中葡萄糖消耗缓慢,一直到24h也没有消耗完。
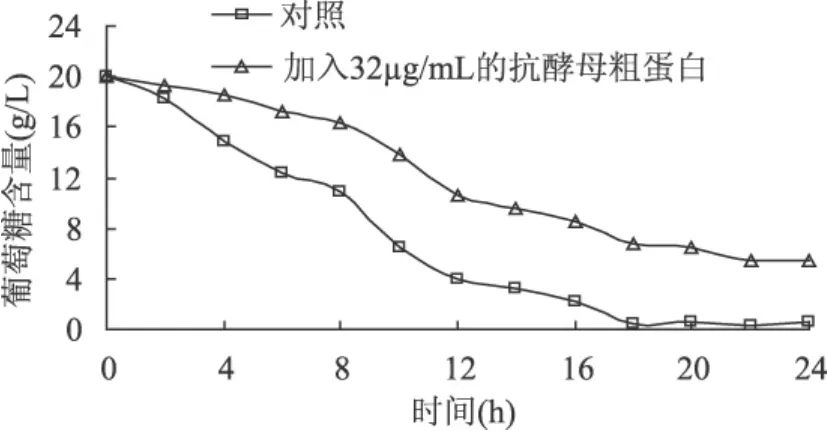
图5 抗酵母蛋白对糖代谢的影响
2.3.2 抗酵母粗蛋白对发酵液中pH的影响 图6显示了发酵液中pH随时间的变化,在对照组中的pH在0~8h下降迅速,从6下降到2.66;而加入抗酵母粗蛋白的实验组pH则下降缓慢,0~8h pH从6下降到4.52。8h后,溶液的pH变化减慢,对照组的pH基本不变,到24h发酵结束时pH为2.13,实验组到24h发酵结束时pH为3.1。
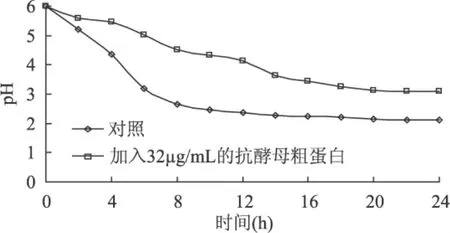
图6 抗酵母活性蛋白对发酵液中pH的变化
2.3.3 不同浓度的抗酵母粗蛋白对酵母生长抑制率的影响 由图7可看出不同浓度的粗蛋白对酵母生长的影响,随着加入的粗蛋白含量的增加,酵母的生长抑制率也在增加,当抗酵母蛋白含量在32!g/mL时,酵母生长抑制率达到了50%,当蛋白含量为50!g/mL时,酵母生长抑制率达到了70%。
3 结论
本文对八种国内外麦芽的抗酵母蛋白进行了含量分析,得出国产Danpi-2的抗酵母粗蛋白含量最高,达到338!g/g,其次是加麦Metcedfe,为322!g/g;同时对抗酵母蛋白的性质进行了分析,得出分子量为10kDa左右的抗酵母粗蛋白能抵抗100℃的高温,加入抗酵母粗蛋白的发酵液中酵母数量大量减少,酵母对葡萄糖的利用下降,pH下降缓慢,当粗蛋白含量达到 32!g/mL时,酵母生长抑制率达到了50%。

图7 不同含量的抗酵母粗蛋白对培养12h后酵母生长抑制率影响
啤酒发酵过程中经常出现的酵母耗糖速度过慢的问题是长期以来困扰啤酒生产企业的主要技术问题之一,其直接导致啤酒发酵周期延长、产量下降,从而增加了啤酒的生产成本。啤酒酵母和麦芽是啤酒生产的两个关键因素,其质量的优劣直接决定着啤酒的风味和成本,很长时间里人们把酵母耗糖速度过慢的问题归结于啤酒酵母的质量,认为是酵母的添加量不足,使用代数过多或者麦汁充氧不足导致酵母活力下降,于是人们通过对现有生产菌种进行改造或筛选新的耗糖速度较快的酵母菌种,或者减少酵母的使用代数的方法试图解决上述问题,但是效果并不是很理想。然而随着啤酒发酵理论和制麦理论的发展,研究者将解决上述问题的注意力逐渐的转移到啤酒的原料——麦芽上,其中抗酵母多肽(简称抗菌肽)的发现标志着向从根本上解决此技术难题迈出了坚实的一步[12-13]。
[1]Boutrot F,Guirao A,Alary R,et al.Wheat non-specific lipid transfer protein genes display a complex pattern of expression in developing seeds[J].BiochimBiophys Acta,2005,1730(2):114-125.
[2]Molina A,Garcia-Olmedo F.Developmental and pathogeninduced expression of three barley genes encoding lipid transfer proteins[J].Plant,1993(4):983-991.
[3]Ara N,Nur MH,Amran MS,et al.In vitro antimicrobial and cytotoxic activities of leaves and flowers extracts from Lippia alba[J].Biol Sci,2009,12(1):87-90.
[4]Garcia-Olmedo F,Molina A,Segura A,et al.The defensive role of nonspecific lipid-transfer proteins in plants[J].Trends Microbiol,1995(3):72-74.
[5]Douliez J P,Michon T,Elmorjani K,et al.Structure,biological and technological functions of lipid-transfer protein and indolines,the major lipid binding proteins from cereal kernels[J].Cereal Sc,2000,32:1-20.
[6]Okada T,Yoshizumi H.A lethal toxic substance for brewery yeast in wheat and barley(Part II).Isolation and some properties of toxic principle[J].Agri Biol Chem,1970,34:1089-1094.
[7]Gorjanovic'S,Sužnjevic'D,Beljanski M,et al.Effects of lipid transfer protein from malting barley on brewers yeast fermentation[J].Inst Brew,2004,110:297-302.
[8]Gorjanovic'S,Spillner E,Beljanski M,et al.Malting Barley Grain Non-specific Lipid-Transfer Protein(ns-LTP):Importance for Grain Protection[J].Inst Brew,2005,111(2):99-104.
[9]Sandra NE,van Nierop,Barry C Axcell,et al.Optimised quantification of the antiyeast activity of different barley malts towards a lager brewing yeast strain[J].Food Microbiology,2008,25:895-901.
[10]Bradford M M.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding[J].Anal Biochem,1976,72:248-254.
[11]郭尧君.蛋白质电泳实验技术[M].北京:科学出版社,2005:58-76.
[12]Wolf-Hall C E.Mold and mycotoxin problems encountered during malting and brewing[J].Int J Food Microbiol,2007,119:89-94.
[13]Wolf-HallC E,SchwarzP B.Mycotoxinsand fermentationbeer production[J].Adv Exp Med Biol,2002,504:217-226.
Character analysis and content comparison of anti-yeast crude protein from several kinds of barley malt
JIANG Xiao-lei,HAN Dan,LI Ming-da,CHEN Xia,ZHAO Rui,ZHAO Chang-xin*
(College of Bio.&Food Tech.,Dalian Institute of Light Industry,Dalian 116034,China)
Out of the eight malt types tested,Danpi-2 barley showed the highest concentration of toxic crude protein with 338!g/g of crude protein,which extracted from malt by sulphuric acid and fractionated by ammonium sulphate precipitation.The character analysis of anti-yeast protein also showed that 10kDa protein tolered temperature as high as 100℃.Adding anti-yeast crude protein into fermentation liquor,the yeast amount decreased remarkably,utilization of glucose decreased and pH decreased slowly.When the concentration of proteins was increased to 32!g/mL,the inhibition ratio also went up by 50%.
barley malt;anti-yeast protein;character analysis
TS210.1
A
1002-0306(2011)02-0112-04
大麦在自然界的生长过程中,不可避免的要遭受自然界中一种或多种病原微生物的侵害,为了抵御这种侵害,大麦自身逐渐形成了各种各样的防御机制[1-2]。一些植物通过分泌一些化学物质来杀灭或抑制某种病原真菌的侵染或生长,有些产生一系列的抗菌蛋白或抗菌多肽,这些抗菌多肽或抗菌蛋白具有广谱抗菌性,到目前为止,一些类型的抗菌蛋白和抗菌肽已经被分离出来,并且有些结构和功能已经研究得比较清楚,如植物或昆虫、哺乳动物防御素、硫堇、橡胶素、类打结素、脂转移蛋白类等[3-5]。1970年,Okada[6]等人从大麦和小麦中分离出了一种抗菌蛋白,分子量在9kDa左右,这种蛋白具有广谱抗菌性,能耐受100℃的高温;2004年和2005年,Gorjanovic',S.[7-8]等人从大麦中分离出来一种分子量在16kDa的蛋白,并指出是一种脂转移蛋白。2008年,Sandra N.E.[9]等人从大麦中粗提出一类具有抗菌作用的多组分蛋白,里面含有Ns-LTP、硫堇、植物防卫素等,具有强烈的抗酵母活性。本文针对国内外几种麦芽进行抗酵母粗蛋白的提取,研究不同品种麦芽的抗酵母粗蛋白含量差别,其对酵母菌生长、糖代谢、pH变化等的影响,为发酵工业尤其是啤酒生产中出现的问题提供理论支持。
2010-02-08 *通讯联系人
姜晓雷(1983-),男,在读硕士研究生,研究方向:国产大麦蛋白分离和制麦工艺。
国家“十一五”国家科技支撑计划重点项目(2007BAK36B01)。
