结合生物学意义的化学计量学数据处理用于微生物代谢指纹分析
2011-09-26吴思,田晶,许卫萍
吴 思, 田 晶, 许 卫 萍
( 1.大连工业大学 生物工程学院, 辽宁 大连 116034;2.大连工业大学 现代教育技术部, 辽宁 大连 116034 )
0 引 言
微生物代谢指纹分析已经被广泛用于微生物的生理、生化和调控等特征的研究,在医学、环境和工业微生物等领域获得了很大成功[1-4]。目前代谢指纹分析中常采用的分析技术包括核磁共振 (NMR)、气相色谱-质谱(GC-MS)、液相色谱-质谱(LC-MS) 以及毛细管电泳-质谱等技术[5-7]。其中GC-MS由于其高灵敏性和具有化合物标准谱库 (NIST) 而得到了更为普遍的应用[7-8]。
GC-MS代谢指纹分析采用的是EI源,因此一次可以产生大量(化合物碎片)数据,其中包括有价值的信息,还包括了许多干扰信息,有些可能是来源于分析方法,有些来自于分析仪器自身,还有一些来源于样品本身。这些不可靠数据的存在常常会给代谢指纹分析带来许多干扰。虽然目前关于代谢指纹分析的原始数据处理和后续的化学计量学分析有许多方法,但是大部分方法更多的是遵循基本的纯统计学原则,而没有更有效地结合生物学意义进行分析[8-11]。为克服上述缺陷,进而能相对合理、准确地利用代谢指纹数据,有效地从庞大的代谢指纹宏数据中分析、提取有价值的信息尤为重要。本研究采用一种结合生物学意义的化学计量学数据处理方法,对不同遗传背景下的6株大肠杆菌的基本代谢特征进行了分析。
1 实 验
1.1 试剂和菌株
N,O-双三甲基硅基三氟乙酰胺(BSTFA+1%TMCS)、甲酸、乙腈、吡啶、甲醇、癸酸,均为分析纯;实验用水为Milli-Q水处理机(MILLPORE 美国公司,密理博上海贸易有限公司)产生的高纯水。
本研究使用6株不同遗传背景的大肠杆菌,详见表1。

表1 实验菌株
1.2 主要仪器
GC-MS QP2010,日本Shimadzu公司;UV-2450紫外分光光度计,日本Shimadzu公司;Biofuge stratos高速冷冻离心机,德国Kendro公司; LRH-150生化培养箱,上海一恒科技;真空冷冻干燥机FREEZONE4.5,美国Labconco公司。
1.3 培养基及培养条件
葡萄糖培养基使用LB培养基,其主要成分为葡萄糖(1.0 g/L),蛋白胨(10.0 g/L),牛肉粉(3.0 g/L)和氯化钠(5.0 g/L)。果糖培养基以同浓度果糖代替培养基中的葡萄糖,其余成分不变。121 ℃高压灭菌后,两种碳源分别以0.22 μm无菌滤膜过滤后以滤液形式加入。培养条件为37 ℃恒温通气摇床(160 r/min)培养。
1.4 细胞干重测定
取10 mL发酵液离心所得沉淀,用生理盐水洗净后在60~100 ℃的烘箱中烘干5 h,称重至恒重(g),即为细胞干重(g)。
1.5 样品处理方法及GC-MS分析方法
在样品处理和淬灭中,快速操作对得到精确的代谢物浓度非常重要,因为很多代谢反应变化都很迅速[12]。因此在对数期将菌液离心并涂于平板上生长2~3 h后,取一定量菌体,放到盛有1.5 mL -20 ℃酸乙腈溶液的器皿内,同时加入70 μL 内标溶液癸酸(1 g/L),而后离心吸取上清液1 mL放入冷冻干燥机中冻干。在冻干后的胞内代谢物中加入吡啶50 μL,超生10 min溶解后,再加入BSTFA+1%TMCS60 μL,75 ℃水浴45 min。104r/min离心10 min后取上清用于GC-MS分析。定量结果以内标[13]及细胞干重进行归一化。
色谱条件:色谱柱,DB-5MS(30 m×0.25 mm×0.25 μm) (J&W, USA);进样1 μL,分流比10∶1,70 ℃ (5 min)―5 ℃/min―280 ℃ (5 min),载气体积流量,He 1.19 mL/min;MS, 进样口280 ℃,离子源200 ℃,传输线280 ℃;离子扫描,m/z33~550,溶剂延迟6 min;检测电压0.9 kV。
2 结果与讨论
2.1 不同碳源条件下变化最显著代谢物的确定
分别在葡萄糖和果糖为唯一碳源培养条件下,对6株大肠杆菌的代谢情况进行GC-MS检测。经XCMS软件分析,分别提取出2 002和1 687个有效碎片离子信息。相应信息经由NIST库和标准品验证分别得到35和24种胞内化合物相对稳定的定性组分,虽然这些化合物在不同条件下都有一定程度的浓度波动,但是,应该有一种化合物表现为在同一培养条件下、不同菌株之间的浓度变化是最显著的。为了分别找到在葡萄糖和果糖培养条件下胞内最显著变化的化合物,作者采用了Tusher等[14]提出的微矩阵显著性分析策略(Significance Analysis of Microarrays ,简称SAM分析),结果表明在葡萄糖为唯一碳源条件下,最显著变化的胞内代谢物是谷氨酰胺,而果糖条件下则是尸胺(图1、2)。
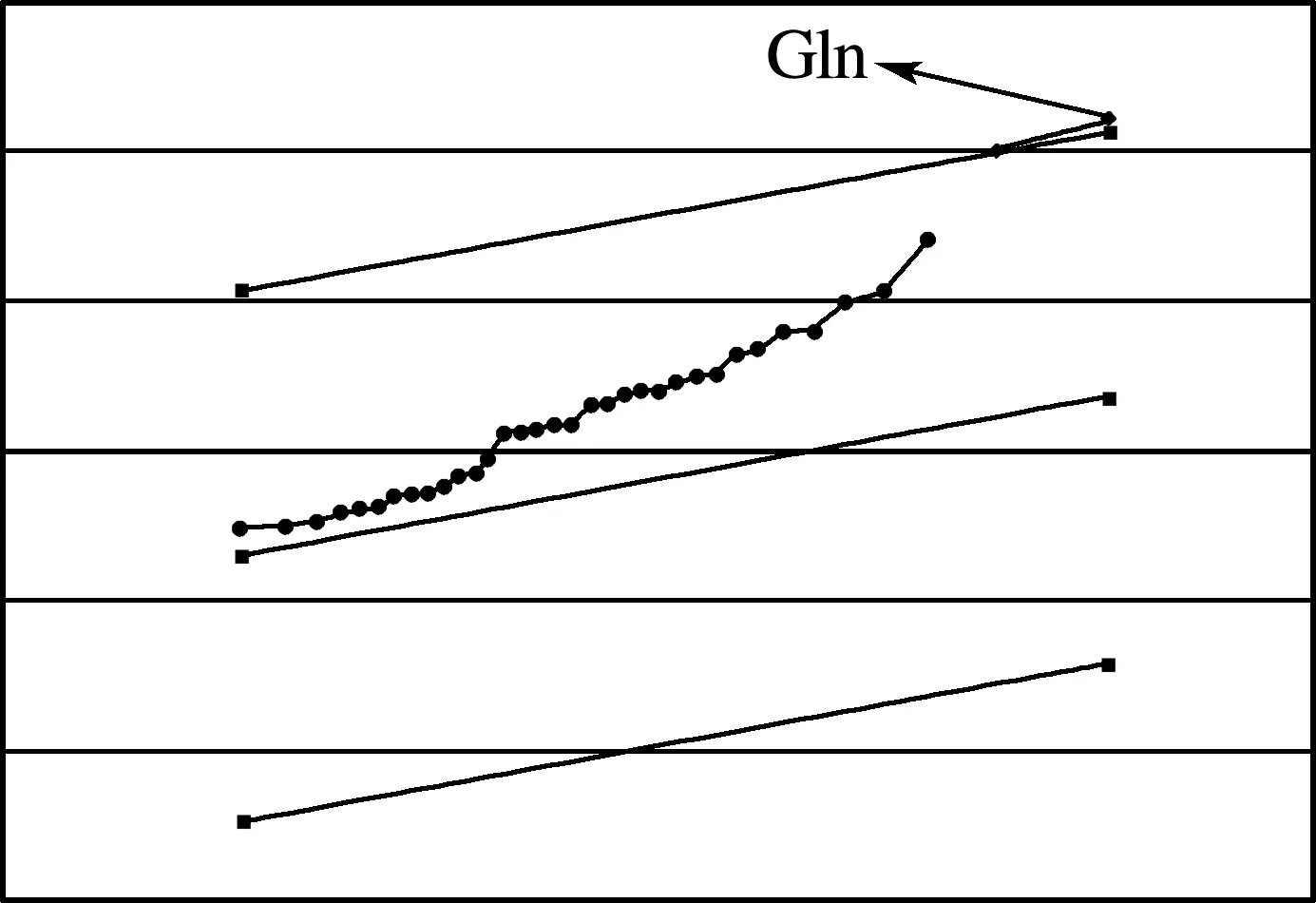
图1 葡萄糖碳源条件下最显著变化化合物的SAM分析结果

图2 果糖碳源条件下最显著变化化合物的SAM分析结果
2.2 不同碳源条件下相关系数较大代谢物的确定
由于微生物细胞内的代谢活动发生在一个庞大的互动网络中,因此,一种代谢物的浓度改变必然会引起其他一些相关代谢物的浓度变化,这些代谢物的协同变化特征则主导了微生物的表型特征,因为代谢特征被认为是最接近于生物表型的反应[13]。为了寻找这部分代谢物,作者又以所发现的最显著变化的代谢物(谷氨酰胺、尸胺)为中心,分别与同条件下检出的其他代谢物进行Pearson相关分析,其中相关系数大于0.6的代谢物(表2)被筛选出来,这些化合物再进行聚类分析。

表2 不同碳源条件下相关系数大于0.6的化合物
2.3 不同碳源条件下聚类分析结果
不同碳源条件下聚类分析结果见图3、4。从图3、4上方聚类线由内向外观察可以发现,6株菌的主体聚类特征比较相似,即无论在何种碳源条件下,菌株arcA、Y和SA都具有相似的聚类特征,而其他3株菌的特征都与这3株菌比较远。
大多情况下遗传特征决定着代谢特征,因此

化合物均经NIST库检索,*为同时经标准品验证
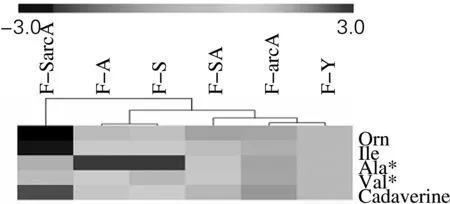
化合物均经NIST库检索,*为同时经标准品验证
在葡萄糖培养条件下具有相似代谢特征的菌株,在果糖条件下也应具有类似的代谢特征。在这3株菌中,arcA基因是与氧供给有关,在该实验中氧气供给充足,所以基因并未发挥其调节作用,生长特性与野生株相差不大,而SA菌株在实际的实验中发现,它有着与arcA较为相似的生长特性。因此至少从arcA、Y和SA这3个样品来说,本研究中对代谢指纹分析数据的处理是恰当而准确的,结合了生物学特征的数据处理过程,还降低了最终参数的个数。其他3株菌聚类特征之间的差异,可能是由其遗传特征的特异性改变所致,作者的前期工作结果也一定程度上提示了环境因素对表型的决定作用。
3 结 论
本研究初步证实了结合一定生物学意义的代谢指纹数据处理技术可以相对有效地提取出有价值的信息,利用该方法能在一定程度上避免其他非相关因素对代谢指纹数据的影响,如果能在更大量的不同菌株测试的基础上对该方法进行验证和调整,将使代谢指纹分析数据得到更合理的应用。
[1] RAAMSDONK L M, TEUSINK B, BROADHURST D, et al. A functional genomics strategy that uses metabolome data to reveal the phenotype of silent mutations [J]. Nature Biotechnology, 2001, 19:45-50.
[2] COMISH-BOWDEN A, CARDENAS M L. Functional genomics:Silent genes given voice[J]. Nature, 2001, 409:571-572.
[3] van der WERF M J, OVERKAMP K M, MUILWIJK B, et al. Comprehensive analysis of the metabolome of Pseudomonas putida S12 grown on different carbon sources[J]. Molecular Biosystems, 2008, 4:315-327.
[4] BRAUER M J, YUAN J, BENNETT B D, et al. Conservation of the metabolomic response to starvation across two divergent microbes[J]. Proceedings of the National Academy of Sciences of USA, 2006, 103:19302-19307.
[5] van der GREEF J, STROOBANT P, van der HEIJDEN R. The role of analytical sciences in medical systems biology[J]. Current Opinion in Chemical Biology, 2004, 8:559-565.
[6] FIEHN O, KOPKA J, DORMANN P, et al. Metabolite profiling for plant functional genomics[J]. Nature Biotechnology, 2000, 8:1157-1161.
[7] KOEK M M, MUILWIJK B, van der WERF M J, et al. Microbial metabolomics with gas chromatography/mass spectrometry[J]. Analytical Chemistry, 2006, 78:1272-1281.
[8] TIAN Jing, SANG Ping, GAO Peng, et al. Optimization of a GC-MS metabolic fingerprint method and its application in characterizing engineered bacterial metabolic shift[J]. Journal of Separation Science, 2009, 32:2281-2288.
[9] VIEITES J M, GUAZZARONI M E, BELOQUI A, et al. Metagenomics approaches in systems microbiology[J]. FEMS Microbiology Reviews, 2009, 33:236-255.
[10] MASHEGO M R, RUMBOLD K, de MEY M, et al. Microbial metabolomics:past, present and future methodologies[J]. Biotechnology Letters, 2007, 29:1-16.
[11] van der WERF M J, JELLEMA R H, HANKEMEIER T. Microbial metabolomics:replacing trial-and-error by the unbiased selection and ranking of targets[J]. Journal of Industrial Microbiology and Biotechnology, 2005, 32:234-252.
[12] BUCHHOLZ A, HURLEBAUS J, WANDEREY C, et al. Metabolomics:quantification of intracellular metabolite dynamics[J]. Biomolecular Engineering, 2002, 19(1):5-15.
[13] TIAN J, SHI C, GAO P, et al. Phenotype differentiation of threeE.colistrains by GC-FID and GC-MS based metabolomics[J]. The Journal of Chromatography B, 2008, 871(2):220-226.
[14] TUSHER V G, TIBISHIRANI R, CHU G. Significance analysis of microarrays applied to the ionizing radiation response[J]. Proceedings of the National Academy of Sciences of USA, 2001, 98:5116-5121.
