抑癌基因RIZ1的研究进展
2011-08-17崔元涛综述张鹏审校
崔元涛综述,张鹏审校
(天津医科大学总医院心胸外科,天津300052)
视网膜母细胞瘤蛋白结合锌指结构基因(retinoblastoma protein-interacting zinc finger gene,RIZ)是由Buyse等[1]应用Rb探针在对可与Rb结合的蛋白质进行功能性筛选时分离出来的,RIZ基因由于转录起始位点的不同可表达RIZ1以及RIZ2两种蛋白质,RIZ1基因属于核组蛋白甲基转移酶超家族[2],是新近发现的肿瘤抑制基因。现就RIZ1基因的定位、结构、功能以及与肿瘤的关系做一综述。
1 RIZ基因定位和结构
Buyse等[1]通过原位杂交荧光技术分析得知RIZ基因定位于人染色体1p36上。RIZ基因能够表达280 kDa的RIZ1及其替换蛋白产物250 kDa的RIZ2,RIZ1含有 PR-domain,RIZ2 PR-domain 缺失,两者其余序列均相同(图1)。人的RIZ1 cDNA全长为7943kb,由10个外显子编码。RIZ1cDNA的大部分由第8外显子编码,PR domain由第4~6外显子编码,RIZ1除含有PR-domain外,尚含有8个锌指结构序列,一个与EIA同源的区,一个GTP motif,一个SH3motif[3]。RIZ2则由5′端临近第5外显子的内部启动子编码,RIZ2内部启动子具有看家基因TATA-less GC-rich的典型特征[4]。
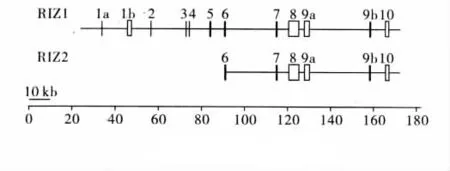
图1 RIZ基因结构图
2 RIZ1的功能
RIZ1含有的PR-domain被称为PRDI-BF1-RIZ1同源区,含有100多个氨基酸,它的生物学功能是蛋白结合面(protein-binding-interface),介导蛋白-蛋白相互作用,对染色体结构的稳定性起重要作用,可调控染色质的基因表达[5]。编码PR-domain的基因家族在肿瘤发生中有着特殊的阴阳作用机制,PR-domain存在或不存在时表达的蛋白产物不同,当一种产物在癌细胞中存在或过表达时,另一种产物则不表达,这两种产物数量不平衡是基因失活的早期变化,是导致肿瘤发生的主要机制之一[6]。Rb是目前最受重视的肿瘤抑制基因之一,作为Rb结合蛋白,RIZ1的基因产物可以与Rb DNA结合,抑制其转录,其编码的RIZ1蛋白与Rb蛋白结合,参与Rb途径发挥抑癌作用[7],此外RIZ1蛋白能导致肿瘤细胞G2/M期阻滞并诱导细胞凋亡[8]。
3 RIZ1基因与肿瘤
RIZ1可以抑制肿瘤的生长,RIZ1基因表达沉默可导致多种肿瘤的发生。在动物实验中Jiang等[9]分别将AdRIZ1病毒和AdGFP病毒感染裸鼠,发现AdRIZ1组6/6在5周的观察期内未发现肿瘤生长,而在AdGFP组4/6的裸鼠在2周后便发现肉眼可见的肿瘤,并呈进行性生长,6/6在3~5周的观察期内均发现肿瘤生长。Steele-Perkins等[7]发现RIZ1基因敲除的鼠模型具有肿瘤易感性,会患弥漫性B细胞淋巴瘤(diffuse large B cell lymphoma,DLBI)和胃癌等多种肿瘤。在细胞实验中Jiang等[9]用腺病毒介导的RIZ1感染肝癌细胞系HuH2,发现感染后的细胞生长缓慢,且在72 h出现死亡,而用单纯腺病毒感染的肝癌细胞无明显的细胞死亡现象。通过DNA histogram分析发现,实验组48 h后出现细胞G2/M期阻滞,并在72 h出现细胞凋亡现象,凋亡程度与感染AdRIZ1的量成正相关。Liusheng等[8]用腺病毒介导的RIZ1转染乳腺癌细胞系UTA6,采取TUNEL分析发现转染后的乳腺癌细胞G2/M期阻滞、细胞发生凋亡。余卫平等[10]将含RIZ1全长cDNA的重组质粒pRIZ1RH转入髓性白血病细胞株AML193和K562,于基因导入24 h后应用FCM测细胞凋亡率同样得出RIZ1的转录能够诱导髓性白血病细胞株凋亡的结论。
RIZ1基因表达下降或缺失发生在多种人类肿瘤中。遗传机制和表观遗传机制均可导致RIZ1基因的失活。遗传机制改变主要指基因突变、缺失和染色体重排等引起的基因序列及其表达产物的改变,RIZ1可通过染色体不稳定性(chromosoimo insta-bility,CIN)和微卫星不稳定性(microsatellite instability,MIN)等途径而失活,并且发生移码突变、点突变、杂合缺失等多种突变印证了这种机制。RIZ基因定位的染色体1p36与多种肿瘤如乳腺癌、卵巢癌、肝癌、结直肠癌、慢性髓性白血病、黑色素瘤、嗜铬细胞瘤、成神经细胞瘤等相关[11]。Liusheng等[8]采取RT-PCR的方法分析乳腺癌细胞系、肺癌细胞系、神经母细胞瘤细胞系以及骨肉瘤细胞系中均发现RIZ1低表达或表达缺失。以后研究陆续发现RIZ1基因表达沉默很普遍的出现在如肝癌、乳腺癌、结肠癌、神经母细胞瘤、肺癌、黑素瘤、骨肉瘤等多种人类原发性肿瘤之中,而这些肿瘤中常存在lp36杂合性缺失佐证了RIZ1基因通过染色体不稳定性的途径失活。RIZ1基因失活的另外一种途径是微卫星不稳定性,研究发现在有微卫星不稳定性的多种人类肿瘤组织、肿瘤细胞系中存在RIZ1移码突变,Pan等[12]应用DHPLC和DNA测序发现11.4%(8/70)的胃癌标本中在BAT25和BAT26存在基因不稳定性,其中7例两者均存在不稳定性,4例(57.1%)存在RIZ1基因移码突变,提示在MSI的胃癌患者中,RIZ1基因的移码突变在失活中起了重要作用。Piao等[13]用PCR-SSCP方法检测除发现48%(19/40)的胃癌组织存在RIZ1基因移码突变外,还发现26%(14/51)的大肠癌组织存在RIZ1基因移码突变,33%(6/18)的子宫内膜癌组织存在RIZ1基因移码突变,54%(7/13)MSI(+)的肿瘤细胞株中存在RIZ1基因移码突变,以上研究均说明了RIZ1基因可通过微卫星不稳定性的途径失活。
遗传学上抑癌基因的突变与肿瘤的发生有着密切的关系已经得到公认,然而近年来随着对肿瘤认识的深入,人们发现DNA序列以外的调控机制 (即表观遗传学)异常在肿瘤的发生、发展过程中也起到非常重要的作用[14]。表观遗传学机制包括:DNA甲基化异常,组蛋白修饰,非编码RNAs(包括microRNA),染色质重塑等,这几个方面都与肿瘤的发生有关[15]。其中DNA甲基化(methylation)是目前研究得最清楚、也是最重要的表观遗传修饰形式[16]。发生在抑癌基因启动子及附近区域的DNA甲基化可直接阻碍转录因子 AP-2、c-Myc、CREB、E2F 和 NFκ-B 与启动子结合,从而使抑癌基因不能转录或转录水平降低[17],同时基因5′端调控序列甲基化后能与特定甲基化CpG序列结合蛋白 (methyl CpG binding proteins,MBPs)结合,间接地阻止转录因子与启动子形成转录复合体[18]。此外,甲基化还能改变染色质的构象,使染色质凝缩成非活性的高级结构[19]。因此,抑癌基因启动子区DNA高甲基化可导致该基因的失活。大量研究表明DNA启动子区CpG岛甲基化也是RIZ1基因表达沉默的最常见原因,Du等[20]在实验中克隆了RIZ1启动子,发现它有典型的CpG岛结构,RIZ1启动子的高度甲基化是mRNA不表达的常见原因。蔡惠兰等[21]采用甲基化特异PCR(methylation specific PCR,MSP)的方法研究发现,25.8%(8/31)的卵巢癌组织及40%(2/5)的卵巢癌细胞系中RIZ1基因启动子区存在甲基化,而且这些存在甲基化的卵巢癌组织、卵巢癌细胞系中RIZ1基因的mRNA表达缺失或下降。Du等[20]采用MSP的方法分析 62.5%(20/32)的肝癌组织,44%(11/25)的乳腺癌组织中RIZ1基因启动子区存在甲基化,用DNA甲基转移酶抑制剂5-杂氮-2-脱氧胞苷(5-Aza-CdR)处理肝癌细胞系HepG2和Huh1,结肠癌细胞系SW48,用RT-PCR分析发现RIZ1可重新表达。方娟娟等[22]应用甲基化特异性PCR法检测人乳腺癌组织和相应癌旁组织及乳腺良性肿瘤组织中RIZ1启动子甲基化状态,34例乳腺癌标本中RIZ1启动子甲基化阳性率为44.1%(15/34)。RIZ1启动子甲基化标本中的RIZ1 mRNA表达水平低于未甲基化标本者,两者差异具有统计学意义(P<0.05),RIZ1表达降低与其启动子高甲基化状态之间有相关性(P<0.05)。王芳等[23]用5-Aza-CdR处理胃癌细胞系BGC82也观察到RIZ1基因启动子区域高甲基化发生逆转从而使基因表达上调。
4 展望
尽管肿瘤的分子发病机制仍然是个正在探索的领域,但对肿瘤的发生与抑癌基因失活密切相关已达成共识。大量研究表明抑癌基因RIZ1的失活可导致多种肿瘤的发生,RIZ1启动子区CpG岛高度甲基化是其失活主要机制,并在癌发生过程中起重要作用。许多肿瘤细胞株RIZ1启动子CpG岛高度甲基化导致的基因表达缺失,经5-Aza-CdR处理后甲基化可得到逆转从而使其表达上调。启动子区CpG岛高甲基化国外已有相关试剂盒用于支气管肺泡灌洗液、淋巴液、痰液、尿液等相关恶性肿瘤的检测,临床上5-Aza-CdR已经应用于骨髓增生异常综合征、急性髓性白血病等血液系统疾病的治疗。因此,进一步研究RIZ1可为将来以此作为目的基因,进行肿瘤的早期诊断和基因靶向治疗创造条件。
[1]Buyse IM,Shao G,Huang S,et al.The retinoblastoma protein binds to RIZ,a zinc-finger protein that shares an epitope with the adenovirus E1A protein[J].Proc Natl Acad Sci USA,1995,92(10):4467
[2]Keun-Cheol K,Geng I,Shi H.Inactivation of a histone methyl transferase by mutations in human cancers[J].Cancer Res,2003,63(22):7619
[3]Buyse IM,Takahashi E,Huang S,et al.Physical mapping of the retinoblastoma interacting zinc finger gene RIZ to D1S228 on chromosome 1p36[J].Genomics,1996,34(1):119
[4]Liu L,Shao G,Steele Perkins G,et al.The tetinoblastoma interacting zinc finger gene RIZ produces a PR domain-lacking product through an intermal promoter[J].Biol Chem,1997,272(5):2984
[5]Huang S,Shao G,Li M,et al.The PR domain of the Rb-binding zinc finger p rotein RIZ1 is a protein binding interface and is related to the SET domain functioning in chromatin-mediated gene expression[J].Biol Sci,1998,273(26):15933
[6]Jing GL,Liu LM,Buyse IM,et al.The yin-yang of PR-domain family genes in tumorigenesis[J].Histol Histopathol,2000,15(1):109
[7]Steele-Perkins P,Wei F,Yang XH,et a1.Tumor formation and inactivation of RIZ1,an Rb—binding member of a nuclear proteinmethyhransferase superfamily[J].Genes Dev,2001,l5(17):2250
[8]He LS,Yu JX,Liu LM,et al.RIZ1,but not the alternative RIZ2 product of the same gene,is underexpressed in breast cancer,and forced RIZ1 expression causes G2-M cell cycle arrest and/or apoptosis[J].Cancer Res,1998,58(19):4238
[9]Jiang G1,Liu LM,Huang S,et al.Decreased RIZ1 expression but not RIZ2 in hepatoma and suppression of hepatoma tumorigenicity by RIZ1[J].Int J Cancer,1999,83(4):541
[10]余卫平,方娟娟,陈宝安,等.RIZ1表达对髓性白血病细胞株凋亡的影响[J].中国癌症杂志,2007,17(1):65
[11]Huang S.The retinoblastoma protein-interacting zinc finger gene RIZ in 1P36-linked cancers[J].Front Biosci,1999,4:D528
[12]Pan KF,Lu YY,Liu YY,et al.Detection of framshift mutation of RIZ in gastric cancer with microsatellite instability[J].World Gatrolenterol,2004,10(18):2719
[13]Piao Z,Hamdick R,Peter B,et al.Frequent frameshift mutations of RIZ in sporadic gastrointestinal and endomatrial carcinomas with microsatellite instability[J].Cancer Res,2000,60(17):4701
[14]Bird A.Perceptions of epigenetics[J].Nature,2007,447(7143):396
[15]Jaenisch R,Bird A.Epigenetic regulation of gene expression:how the genome integrates intrinsic and environmental signals[J].Nature Reviews Genetics,2003,33(4):245
[16]孙树汉.肿瘤的表观遗传学研究[J].中国肿瘤生物治疗杂志,2008,15(1):8
[17]Belinsky SA.Gene promoter hypermethylation as a biomarker in lung cancer[J].Nat Rev Cancer,2004,4(9):707
[18]Costeuo J,Frnhwald M.Aberrant CpG-island methylatin has nonrandom and tumours-type-specific patterns[J].Nature Genetics,2000,24(2):132
[19]Rountree MR,Bachman KE,Herman JG,et al.DNA methylation,chromatin inheritance,and cancer[J].Oncogene,2001,20(24):3156
[20]Du Y,Carling T,Fang W,et al.Hypermethylation in human cancers of the RIZ1 tumor suppressor gene,a member of a histone/protein methylt ransferase superfamily[J].Cancer Res,2001,61(22):8094
[21]蔡惠兰,钱颖,贺晓琪.等.卵巢上皮性癌中RIZ1基因表达缺陷的意义及其与甲基化的关系 [J].武汉大学学报(医学版),2008,29(4):430
[22]方娟娟,余卫平,安艳丽.乳腺癌中RIZ1基因启动子甲基化状态的探讨[J].中国肿瘤临床,2007,32(12):664
[23]王芳,卢启明.5-杂氮-2-脱氧胞苷对BGC823胃癌细胞株RIZ1基因去甲基化转录调控[J].第四军医大学学报,2008,29(14):1275
