井冈山猴头杜鹃群落特征的研究
2011-07-14邓贤兰吴杨刘玉成钟娟
邓贤兰, 吴杨, 刘玉成, 钟娟
1. 江西省井冈山大学生命科学学院,江西 吉安 343009;2. 西南大学生命科学学院,重庆 北碚 400715
猴头杜鹃(Rhododendron simiarum) 隶属于杜鹃花科(Ericaceae)杜鹃属(Rhododendron),为我国特有的常绿小乔木,分布于我国江西、湖南、福建、浙江、广东和广西等省,生长在海拔900 m以上生态环境条件严酷的山巅、山脊及其坡面上,是亚热带山地矮林的建群种之一[1],在维护林地环境、涵养水源、保护生态平衡等方面起着重要的作用。目前有关猴头杜鹃的研究主要集中在其生长特性[2]、木纤维形态特征和化学成分[3]、群落学特点[1,4]、种群结构和分布格局[5,6,7]等方面,而有关井冈山猴头杜鹃群落的研究未见报道。因此本文通过实地调查,对井冈山猴头杜鹃群落的物种组成、区系成分、群落结构和物种多样性等进行了分析研究,为深入了解其群落特征、科学管理和利用猴头杜鹃林、有效保护种质资源和生物多样性等提供科学的理论依据。
1 研究区概况
井冈山地处南岭山地罗霄山脉中段,在江西境内的一组山体。位于 26°27′~26°40′N,113°39′~114°23′E,整个山体呈东北—西南走向,境内山势高峻、群山层叠、峰丘相间、岭谷夹持、地形复杂。最高海拔1841 m,最低海拔202 m,相对高差1639 m,土壤以山地黄壤为主,土层厚度一般在50~80 cm,土质疏松、肥沃、湿润。井冈山属亚热带温暖湿润气候区,年平均气温14.2 ℃,年降雨量1856.2 mm,由于井冈山地处欧亚大陆东南部南亚热带与中亚热带的过渡地带,长期以来受东亚季风气候的影响,冬温夏凉、春暖秋爽,加之井冈山地势崎岖、多变,形成了局部气候条件的差异,故孕育了大面积的森林植被[8]。
2 研究方法
2.1 样地调查
在猴头杜鹃分布较集中的井冈山笔架山和大井,采用典型取样方法,设置5个20 m×20 m的样地,每个样地划分为4个10 m×10 m的样方,每个样方内设置4个5 m×5 m的灌木样方,4个2 m×2 m的草本样方。调查记录乔木层树种(树高﹥3 m,围径﹥10 cm)的植物名称、围径、高度、冠幅,草本植物与灌木的植物名称、株数(株丛)、高度、盖度等。同时测定海拔、坡度、坡向和土壤深度等环境因子(表1)。
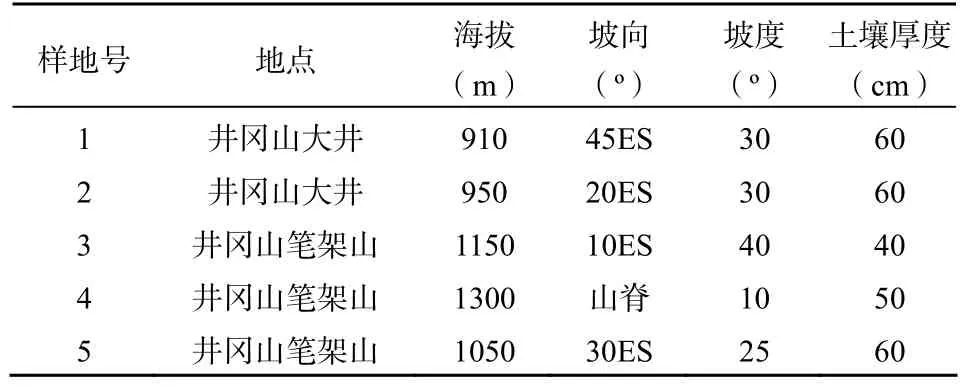
表1 井冈山猴头杜鹃群落样地环境特征Table 1 The environment conditions of the plots in Rhododendron simiarum communities in Jinggang Mountain
2.2 数据处理
分别计算各样地乔木层、灌木层、草本层的重要值,计算公式为
(1)乔木层的重要值:IV=(相对频度﹢相对密度﹢相对显著度)/3;
(2)灌木层及草本层的重要值:IV=(相对频度﹢相对密度)/2;
群落物种多样性指数本文选用应用较广泛的 4种测度方法[9]:
(1)丰富度(S),指样地内所有物种数目。
(3)Shannon-wiener指数:
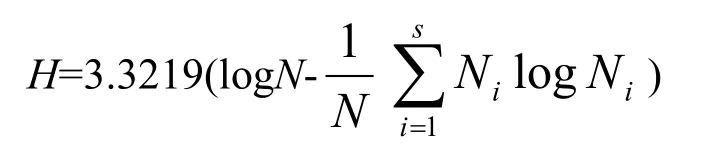
式中,S为样地中物种的种数;Ni为种i的重要值;N为样地中所有物种的重要值之和;Pi为种i的相对重要值,Pi=Ni/N
3 结果与分析
3.1 群落物种组成成分分析
根据井冈山猴头杜鹃植物群落样地调查资料中所记载的植物,统计该群落有维管植物 50种,隶属于30科43属,其中蕨类植物6种,隶属于5科6属,裸子植物4科4属4种,被子植物21科33属40种(表2)。从其生活型来看,木本植物占绝对优势,共有38种,占总种数的76.00%,草本植物12种,占总种数的24.00%。在木本植物中,乔木23种,灌木(含乔木幼树)23种,藤本植物3种。其中常绿乔灌木 26种,占木本植物总数的68.42%。由于常绿木本植物占有较大比例, 可以在一定程度上反映出猴头杜鹃群落植物在系统演化上处于较为古老的位置。

表2 井冈山猴头杜鹃群落物种组成情况Table 2 The species composition in Rhododendron simiarum communities in Jinggang Mountain
3.2 群落植物区系成分统计分析
井冈山的常绿阔叶林群落不仅具有亚热带植被的一般特征, 而且具有本地区常绿阔叶林群落独特的特征。根据中国的植物区系分区[10], 井冈山在中国植物区系分区上属于泛北极植物区、中国-日本植物亚区的中国南部亚热带湿润森林植物区系。分布于井冈山的猴头杜鹃群落是组成井冈山山顶常绿阔叶矮林的重要群系。
根据吴征镒、李锡文[11,12]等学者的观点,并参照等姚振生[13]、邓贤兰[14]的研究报道,将井冈山猴头杜鹃群落维管植物30科43属进行分类(表3)。从表3可以看出,井冈山猴头杜鹃群落30科,分属于5个分布区类型3个变型;43属,分属于9个分布区类型4个变型。
3.3 群落植物区系的主要特征
3.3.1 区系地理成分复杂,热带性成分占优势
井冈山猴头杜鹃群落物种组成中,地理成分复杂。从科级统计来看,具有5个分布区类型3个变型。除世界分布外,热带性科(第 2~7项)有 15科,占总科数的62.50%,其中以泛热带分布科为主,有 11科,占总科数的 45.83%,如里白科(Gleicheniaceae),樟科(Lauraceae)、壳斗科(Fagaceae)、山茶科(Camelliaceae)、杜英科(Elaeocarpaceae)等;温带性科(第 8~14项)有 9科,占总科数的37.50%,其中以北温带分布科为主,5科,占总科数的20.83%,如柏科(Cupressaceae)、松科(Pinaceae)、杜鹃花科(Ericaceae)、槭树科(Aceraceae) 等;世界分布科有 6科,为石松科(Lycopodiaceae)、蝶形花科(Papilionaceae)和莎草科(Cyperaceae)等。在组成猴头杜鹃群落的43个属中,除温带亚洲分布、地中海区、西亚至中亚分布和中亚分布外,其它分布类型或其变型均有。除世界分布属外,第 2~7类热带性属 28个,占总属数的71.79%,其中泛热带分布属最多,有 14属,如樟属(Cinnamomum)、杜英属(Elaeocarpus)、山矾属(Symplocos)、菝葜属(Smilax)等;其次为热带亚洲分布属及其变型,为 10属,如福建柏属(Fokienia)、含笑属(Michelia)、山胡椒属(Lindera)、润楠属(Machilus)、木荷属(Schima)等;温带性分布属(第8-14类)共9属,占总属数的23.08%,其中以北温带分布属最多,有 5属,如贯众属(Cyrtomium)、松属(Pinus)、杜鹃属、水青冈属(Fagus)等;东亚和北美间断分布属2属,为枫香树属(Liquidambar)和栲属(Castanopsis);东亚分布属 2属,占总属数的 5.13%,为五加属(Acanthopanax)和吊钟花属(Enkianthus)。中国特有属 2属,占总属数的 5.13%,为杉木属(Cunninghamia)和白豆杉属(Pseudotaxus)。在猴头杜鹃群落的区系组成中,热带性成分占绝对优势,这与井冈山其它常绿阔叶林区系特征[15]相一致。虽然群落区系组成中以热带成分占优势, 但温带性成分也有一定的比例,这反映出井冈山猴头杜鹃群落植物区系地理成分的复杂性。
3.3.2 植物区系起源古老
井冈山植物起源古老,素有“第三纪型森林”之称[8]。在井冈山猴头杜鹃群落中,其古老和原始性状,首先表现在组成该群落乔木层的建群种和灌木层的优势种,在植物系统演化中就有原始的科属,这些树种都是在白垩纪就已经发生,而且一直繁衍兴盛到现代,如柏科的福建柏属、壳斗科的栲属,樟科的润楠属、樟属、山胡椒属,山茶科的木荷属,木兰科的含笑属等的乔、灌木;其次组成该群落的蕨类植物中也有古老原始的植物,如中生代已存在的里白科的芒萁(Dicranopteris dichotoma) 和见于新生代第三纪的乌毛蕨科的狗脊蕨(Woodwardia japonica)等。
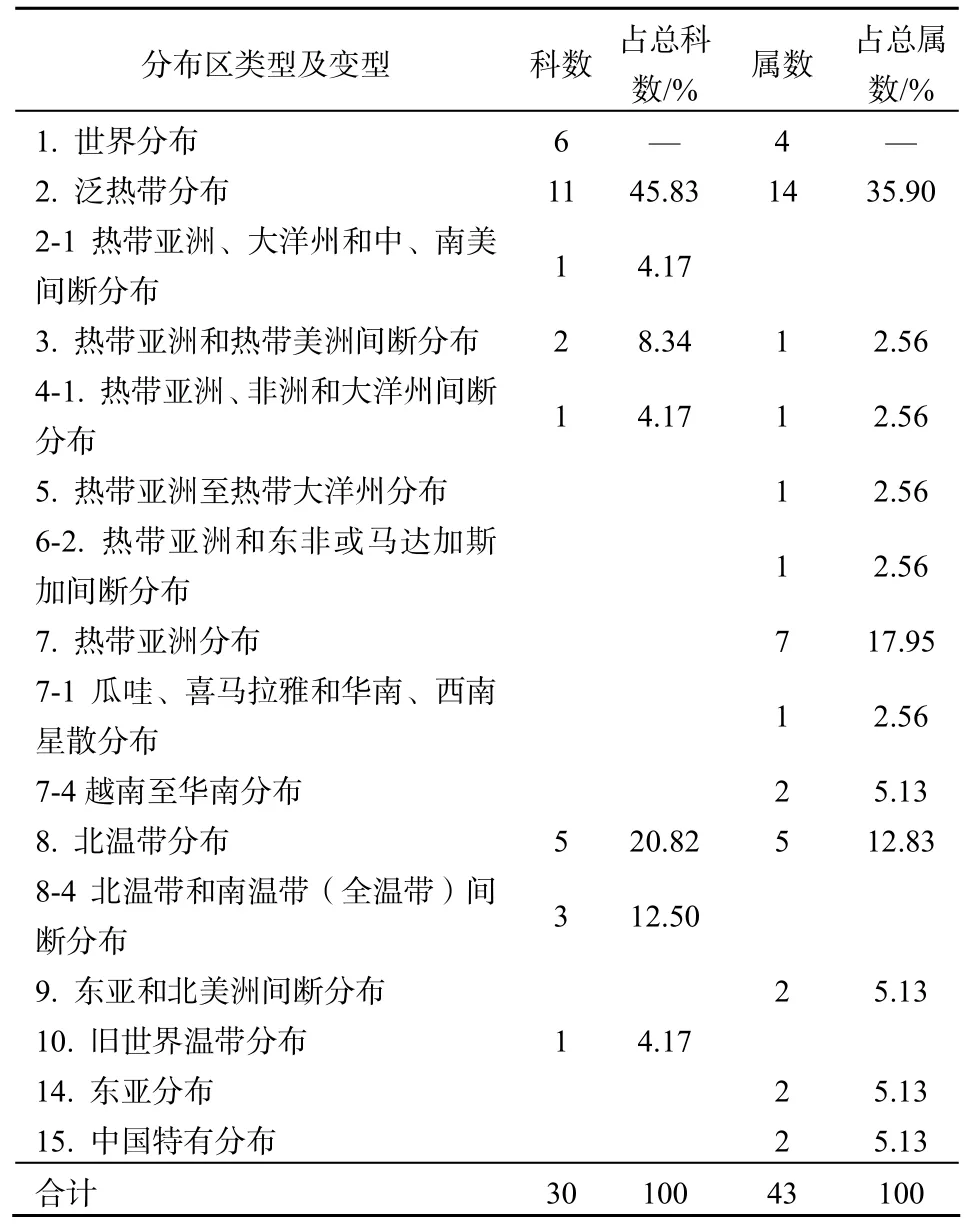
表3 井冈山猴头杜鹃群落维管植物科、属分布区类型Table 3 The Family and Genus distribution types of vascular plants in Rhododendron simiarum communities in Jinggang Mountain
3.4 群落结构
猴头杜鹃是我国亚热带山顶矮林植被中分布最广、面积最大的群系[1],在井冈山主要集中分布在笔架山、大井等地海拔900~1400 m之间的山坡和山脊上,林冠整齐,长势较繁茂。猴头杜鹃、甜槠(Castanopsis eyreiTutch)、福建柏(Fokienia hodginsii)、台湾松(Pinus taiwanensis)、薯豆(Elaeocarpus japonicus)和白豆杉(Pseudotaxus chienii)是组成猴头群落的建群种,对群落的结构和群落环境的形成起着重要的作用。其中猴头杜鹃、福建柏、甜槠从幼树到大树均有,为多世代植物组成。
调查群落所处坡向为东南方向或山脊,坡度10°~40°,土壤为山地黄壤,土层厚度40~60 cm左右。群落由乔木层、灌木层和草本层组成。乔木层的郁闭度为0.85左右,乔木层层高4 m~11 m,主要由猴头杜鹃、福建柏、甜槠、台湾松、薯豆、马银花(Rhododendron ovatum)、枫香(Liquidambar formosana)组成。灌木层高0.2 m~3 m,盖度20%~50%。主要树种有猴头杜鹃、柃木(Eurya japonica)、细叶香桂(Cinnamomum chingii)、甜槠、福建柏、深山含笑(Michelia maudia)等。草本层盖度5%~25%,主要有求米草(Oplismenusundulatifolius)、芒尖苔草(Carex doniana)、狗脊蕨、贯众(Cyrtomium fortunei)等。层外藤本植物有菝葜(Smilax china)和木通(Stephanotis chunii)等。
3.5 群落物种多样性分析
5个样地均为常绿阔叶林或针阔混交林,通过计算得到5个样地不同生活型植物的多样性指数值并作图1以及不同层次的多样性指数值及其变异系数(表 4)。从图 1可知井冈山猴头杜鹃群落,除Pielou指数较高外,Simpson指数、Shannon-wiener指数和物种丰富度指数均较低,并且各样地间的多样性指数测值变幅除Pielou指数较小外,其它3种指数变幅均较大;测度群落物种多样性时,各种多样性指数基本表现出相同的趋势,但因群落类型、生境条件的不同和人为因子的影响,各样地间物种多样性有一定的差异,这反映了多样性指数能反映群落的状况。如样地4,处在笔架山海拔1300 m的山脊,乔木层优势种单一,为猴头杜鹃,土层脊薄、干燥,加上每年观赏的游客较多,人为影响较大,所以其多样性指数较低;样地1和样地2,位于大井海拔九百多米的山坡,都为针阔混交林,群落内土层较厚、湿润,人为干扰较轻,物种多样性指数较高。这与“生境条件优越,人为干扰轻,则多样性指数就高”这条规律[16,17]相一致。
井冈山猴头杜鹃群落的物种多样性指数随海拔高度变化的趋势不十分明显,这与植物群落物种多样性指数在海拔剃度上未表现出明显的规律性[18]是一致的。但山地分布区域的环境条件、山地的相对高度、人为干扰的程度、不同海拔的群落类型等都有可能对植物群落的物种多样性指数在海拔剃度上的分布产生影响[19,20]。虽然样地3、4、5都位于笔架山,但样地3、5所处的海拔与样地4相比低150 m~250 m,且都位于阳坡,土层较厚,人为干扰相对较小,物种多样性指数相应高一些。
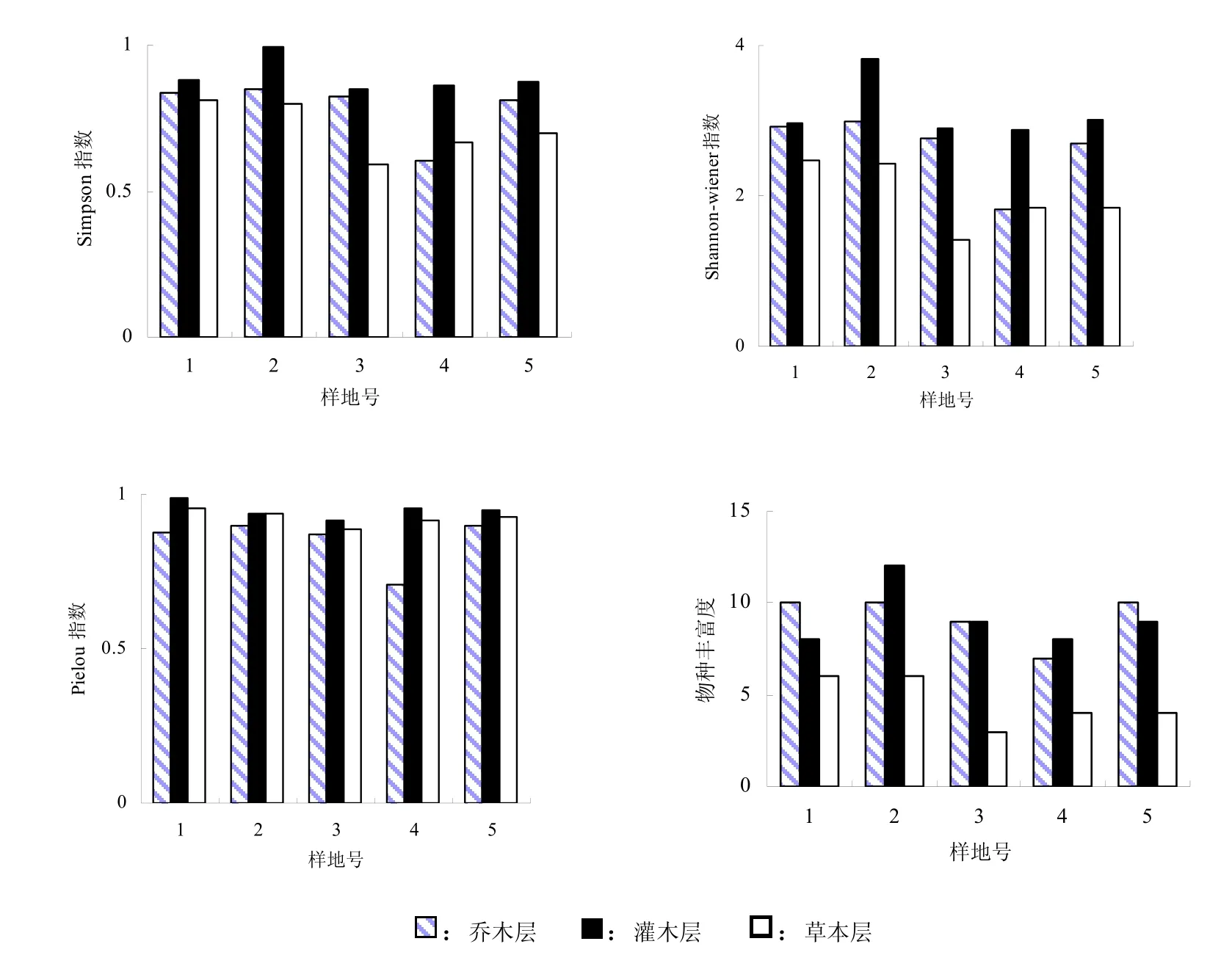
图1 井冈山猴头杜鹃群落物种多样性指数Fig.1 Species diversity index of Rhododendron simiarum communities in Jinggang Mountain
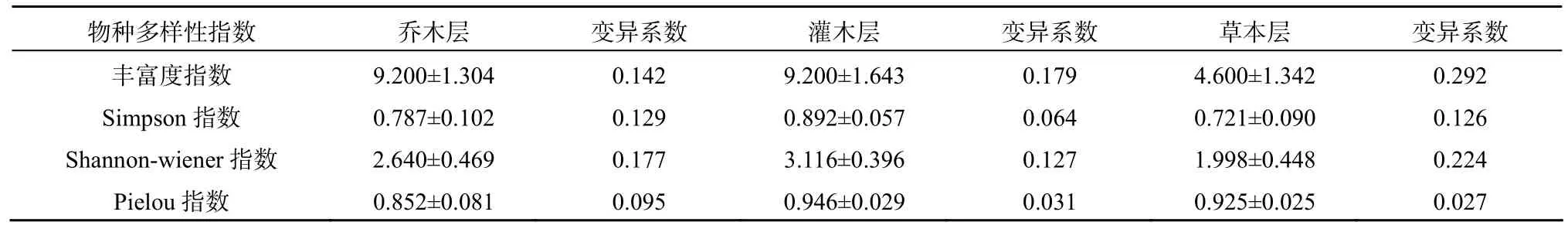
表4 井冈山猴头杜鹃群落不同层次物种多样性指数Table 4 Species diversities of the different layer in the Rhododendron simiarum communities in Jinggang Mountain
群落组成树种的数量、空间配置不同,形成了不同的结构格局,其物种多样性也不同,物种多样性作为测定群落结构水平的指标,可以较好地反映群落的结构[17]。井冈山猴头杜鹃群落的垂直结构可分为乔木层、灌木层和草本层等。中亚热带森林群落中,生物量及所占空间均是乔木层最大,灌木层次之,草本层最小,但物种多样性却表现出不同的趋势。从表4可知,不同层次的物种多样性不同,就各种多样性指数而言,基本表现为灌木层>乔木层>草本层,这是由于乔木层中猴头杜鹃、台湾松、甜槠、薯豆占绝对优势,组成乔木层的种类相对较少,且乔木层优势种猴头杜鹃、甜槠树的个体较集中,其它种类的个体数分散,这样乔木层的物种多样性低,而灌木层不仅有灌木种类,还包括乔木的幼树,因此组成灌木层的种类较多,各个种的个体数分布也较均匀,所以物种多样性较高,在群落内由于乔木层、灌木层植物茂盛,郁闭度和盖度较大,地面光照弱,且林地内枯枝落叶层较厚,致使草本植物稀疏,种类少,因此,对草本植物来说,不仅物种间个体数的分配不均匀,而且物种在群落中的空间分布也不均匀,造成样地间的种类组成、个体数量差异较大,因而草本植物的物种多样性最低,这说明群落的物种多样性能反映群落的组织特征和结构状态。从乔木层、灌木层、草本层的变异系数来看,四种指数在不同层次中并没有表现出一定的规律,如丰富度指数和Shannon-wiener指数的变异系数以草本层最大,而Simpson指数和Pielou指数的变异系数以乔木层最大,灌木层介于两者之间。
群落物种组成和结构发生变化,从而导致群落物种多样性和个体数量的降低,如样地4,位于著名的笔架山“十里杜鹃”林中,每年开花季节游客较多,受人为的干扰较大,因此其各层次的物种数量少,尤其是乔木层只有7个种,且优势种单一,物种种多样性指数低。
4 结论与讨论
(1)井冈山猴头杜鹃群落植物种类较丰富,有维管植物30科43属50种,其中蕨类植物有5科6属6种,裸子植物4科4属4种,被子植物21科33属40种。维管植物科、属、种的数量均比广东石门台保护区猴头杜鹃群落(维管植物47科78属112种)少[4],这与所调查群落面积的大小及分布、群落所处的地理位置和气候条件的不同及人为因素的影响有关;尤其是人为因素的影响。笔架山是井冈山猴头杜鹃林分布最集中的地方,近年来由于笔架山“十里杜鹃”林旅游景点的开发及游客的增加,导致笔架山猴头杜鹃群落均受到一定的人为干扰,尤其是处于“十里杜鹃”林中的猴头群落所受的干扰较大,因此植物种类较少。
(2)井冈山猴头杜鹃群落植物区系起源古老,地理成分复杂,热带性成分占优势,温带性成分占有一定的比例。这一研究结果与邓贤兰[15]关于井冈山栲属群落植物区系研究结果相一致。猴头杜鹃群落系中国中亚热带地区常见的常绿阔叶山顶矮林,属于中国广大中亚热带植物区系的一部分,由于受到井冈山山地地形和中亚热带季风气候的影响,并反映出典型的中亚热带植物区系的特点。
(3)井冈山猴头杜鹃群落为复层异龄结构,猴头杜鹃、甜槠、福建柏和台湾松为该群落的建群种。猴头杜鹃群落是地形顶极和地带性的常绿阔叶林植被类型, 它反映着中国亚热带地区海拔900~2000 m地段的大气候状况, 也反映山顶局部气候因素的影响,猴头杜鹃等山顶矮林的兴衰和动态,将直接或间接地影响其周围的常绿阔叶林、亚热带针叶林以及竹林等, 并且猴头杜鹃群落是研究山顶矮林生态学规律的理想对象和基地[1,6]。因此,对井冈山猴头杜鹃群落特征研究,有助于进一步了解猴头杜鹃群落的生态学特征,从而为井冈山猴头杜鹃资源的保护和可持续利用,以及生态环境的保护提供科学的理论依据。
(4)除Pielou指数外,井冈山猴头杜鹃群落物种多样性指数较低,各样地间多样性指数测值变幅较大;测度猴头杜鹃群落物种多样性时,各种多样性指数基本表现出相同的趋势;总体来看,猴头杜鹃群落物种多样性在群落不同层次的变化规律为:灌木层>乔木层>草本层。井冈山猴头杜鹃群落物种多样性指数较低,这与群落所处的地理位置和气候条件的不同及人为因素的影响有关;尤其是人为因素的影响导致物种多样性指数显著的降低。
猴头杜鹃花大美丽,具有重要的观赏价值。近年来,当地农民为了增加家庭经济效益,将群落中猴头杜鹃幼树甚至大树挖掘出来移栽或盆栽出售,这样导致群落中幼树少,如果不加以管理,在长期更新演替过程中,可能出现“青黄不接”现象,也会导致天然旅游资源的浪费或丧失。因此,应采取有力的措施,合理地利用与保护井冈山的猴头杜鹃,使其资源永续利用,同时更应重视对其所在群落及生境的保护。
[1] 叶居新. 中国的猴头杜鹃矮林[J]. 武汉植物学研究, 1994, 12(2) :170-174.YE Juxin. On theRhododendron simiarumelfin forest of China[J].Journal of Wuhan Botanicl Research, 1994, 12(2): 170-174.
[2] 郑凌峰. 天然猴头杜鹃生长特性研究[J]. 江西农业大学学报, 2000,22(3) : 425-427.ZHENG Linfeng. A Study on the Growth Speciality ofRhododendron simiarumfrom Natural Forest[J]. Acta Agriculturae Universitatis Jiangxiensis, 2000, 22(3) : 425-427.
[3] 郑凌峰, 黄东明, 游生勇, 等. 猴头杜鹃木纤维形态特征和化学成分研究[J ]. 亚热带植物通讯, 2000 , 29(2): 23- 26.ZHENG Linfeng, HUANG dongming, YOU Shengyong. Fiber morphological character and chemical composition of the woodRhododendron simiarum[J]. Subtropical Plant Research Communication,2000, 29(2): 23-26.
[4] 王厚麟, 黄林生, 缪绅裕, 等. 广东石门台保护区猴头杜鹃群落特征[J]. 广西植物, 2008, 28 (1) : 73- 77.WANG Houlin, HUANG Linsheng, MIAO Shenyu. Community characteristics ofRhododendron simiarumin Shimentai Nature Reserve , Guangdong[J]. Guihaia, 2008, 28(1): 73-77.
[5] 金则新, 周荣满, 叶立新. 猴头杜鹃种群结构和分布格局研究[J]. 安徽农业科学, 2006, 34(22) :5860-5862.JIN Zexin, ZHOU Rongman, YE Lixin. Population Structure and Distribution Pattern ofRhododendron simiarumPopulation in Fengyang Mountain Natural Reservation[J]. Journal of Anhui Agricultura Science, 2006, 34(22): 5860-5862.
[6] 肖宜安, 胡文海, 黄族豪,等. 井冈山自然保护区猴头杜鹃花种群结构和动态[J].井冈山学院学报, 2007, 28(10): 5-8.XIAO Yian, HU Wenhai, HUANG Zhuhao. Population structure and dynamics ofRhododendron simiarumin Jinggangshan National Natural Reserve[J]. Journal of Jinggangshan University(Natural Sciences), 2007, 28(10): 5-8.
[7] 何小娟, 何东进, 洪伟, 等. 天宝岩天然猴头杜鹃种群空间分布格局研究[J]. 云南农业大学学报, 2009, 24(5): 734-738.HE Xiaojuan, HE Dongjin, HONG Wei, et al.. Study on SpatialD istr ibution of NaturalRhododendron simiarumPopulations in Tianbaoyan Nature Reserve[J]. Journal of Yunnan Agricultural University, 2009,24(5): 734-738.
[8] 林英. 井冈山自然保护区考察研究[M]. 北京: 新华出版社, 1990,140- 198.
[9] 王祖华, 李瑞霞, 王晓杰, 等. 间伐对杉木人工林林下植被多样性及生物量的影响[J]. 生态环境学报, 2010, 19(12): 2778-2782.WANG Zuhua, LI Xiaoxia, WANG Xiaojie, et al.. Effects of thinning on biomass and species diversity of understory in Chinese fir plantations[J]. Ecology and Environmental Sciences, 2010, 19(12):2778-2782.
[10] 王荷生. 植物区系地理[M ]. 北京: 科学出版社, 1992.
[11] 吴征镒. 中国种子植物属的分布类型[J]. 云南植物所研究, 1991,(4): 1-3.WU Zhengyi. The areal-types of Chinese genera of seed plants[J].Acta Botanica Yunnanica, 1991(supp):1-139.
[12] 李锡文. 中国种子植物区系统计分析[J]. 云南植物研究, 1996,18(4): 363-384.LI Xiwen. Floristic statistics and analyses of seed plants from China[J].Acta Botanica Yunnanica, 1996, 18(4): 363-384.
[13] 姚振生, 杨武亮. 江西药用蕨类植物的地理分布及区系特征[J]. 武汉植物学研究, 1996, 14(13): 207-212.YAO Zhengsheng, YANG Wuliang. The geographical distribution and flora feature of the medicinal Pteridopyhtes in Jiangxi, China[J].Journal of Wuhan Botanicl Research, 996, 14(13): 207-212.
[14] 邓贤兰, 黄彩霞. 井冈山自然保护区药用蕨类植物资源与区系分析[J]. 植物研究, 2002, 22(1): 91-97.DENG Xianlan, HUANG Caixia. Analyses on the flora and studies on the function of medicinal Pteridophytes in Jinggangshan Nature Reserve[J]. Bulletin of Botanical Research, 2002, 22(1): 91-97.
[15] 邓贤兰. 井冈山自然保护区栲属群落区系分析[J]. 武汉植物研究,2003, 21(1): 61-65.DENG Xianlan. Floristic Analysis ofCastanopsisCommunities in Jinggang Mountain[J]. Journal of Wuhan Botanicl Research, 2003,21(1): 61-65.
[16] 贺金生, 陈伟烈, 李凌浩. 中国中亚热带东部常绿阔叶林主要类型的群落多样性特征[J].植物生态学报, 1998, 22(4): 303-311.He Jinsheng, Chen Weilie, Li Linghao. Community diversity of the main types of the evergreen broad-leaved forest in the Eastern part of the Middle Subtropical China[J ] . Acta Phytoecologica Sinica, 1998,22(4): 303-311.
[17] 邓贤兰, 龙婉婉, 许东风, 等. 井冈山自然保护区福建柏群落的研究[J]. 热带亚热带植物学报, 2008, 16 (2) : 128-133.DENG Xianlan, LONG Wanwan, XU Dongfeng, et al... Studies onFokienia hodginsiicommunity in Jinggangshan Nature Research[J ].Journal of Tropical and Subtropical Botany, 2008, 16 (2) : 128-133.
[18] 王燕, 刘苑秋, 杨清培, 等. 江西大岗山常绿阔叶林群落特征研究[J]. 江西农业大学学报, 2009, 31 (6): 1055-1062.WANG Yan, LIU Yuanqiu, YANG Qingpei, et al.. A Study on the Community Characteristics of Evergreen Broad-leaved Forest in agangshanMountains of Jiangxi Province[J]. Acta Agriculturae Universitatis Jiangxiensis, 2009, 31 (6): 1055-1062.
[19] Wilson JB, Lee WG, Mark AF. Species diversity in relation to ultramafie substrate and to altitude in southwestern New zealand[J].Vegetatio, 1990. 86: 15-20.
[20] 邓贤兰, 肖春玲, 刘玉成. 井冈山自然保护区栲属群落物种多样性研究[J]. 广西植物, 2004, 24(1): 7-11.DENG Xianlan, XIAO Chunling, LIU Yucheng. Studies on species diversity ofCastanopsiscommunity in Jinggang Mountain Nature Reserve[J]. Guihaia, 2004, 24(1): 7-11.
