攀枝花苏铁类群的种子植物区系地理研究
2011-06-26郝云庆余志祥杨永琼
郝云庆,莫 旭,余志祥,杨永琼
(1.四川省林业科学研究院,四川 成都 610081;2.四川攀枝花苏铁国家级自然保护区管理局,四川 攀枝花 617000)
苏铁类植物是一类起源古老而孑遗的裸子植物,其起源可追溯至古生代二叠纪[1],在中生代晚三叠纪至早白垩纪最为繁盛,晚白垩纪时逐渐衰退,遂由被子植物所取代,现今仅有少数类群残存下来,成为处于濒危状态的孑遗植物。苏铁科仅含苏铁属一个属,被世界公认为是最为原始的属[2]。攀枝花苏铁Cycas panzhihuaensis是在四川南部金沙江干热河谷首次发现的苏铁属植物,早在1951年植物学家就已经采到攀枝花苏铁的标本,当时误认作篦齿苏铁C.pectinata,后又误作云南苏铁C.siamensis。直到1981年在大量调查研究的基础上才得以正确命名[3]。攀枝花苏铁生长在金沙江干热河谷的特殊环境中,最初发现于攀枝花市附近的巴关河西坡,以后在毗邻的格里坪后山及宁南、德昌、盐源和云南华坪等县的局部地区也相继发现有分布。它的发现不仅表明横断山区目前仍存在有天然苏铁群落,特别是宁南种群的分布把苏铁属植物分布的北界推移到北纬27°11'[4]。但进入上世纪80年代后,苏铁的园艺价值被开发,各地兴起的“苏铁热”诱发了大规模盗挖和贩卖野生苏铁风潮,在交通方便的分布区,攀枝花苏铁基本上被盗挖一空。原本分布于金沙江干热河谷及其支流河谷中的13个攀枝花苏铁自然种群,因为商业采挖已使8个种群灭绝[5]。据后来实地调查,除攀枝花苏铁自然保护区外,其余4个分布点已很难见到野生种群。现在面积仅有1358 hm2的攀枝花苏铁自然保护区,集中了全国一半以上的野生苏铁资源,使之成为了世界迄今为止发现的纬度最高,面积最大,数量最多,分布最集中的野生苏铁种群。
植物区系是指某一地区(或分类单元、植物群落)所有植物的总和,是植物界在一定的自然条件下,特别是在自然历史条件下总和作用、发展、演化的结果[6,7]。因此植物区系的组成中蕴涵着大量历史、地理、生态和系统进化信息。攀枝花苏铁作为地史年代古苏铁植物演化的直接后裔,在地史演变和植物系统发生上都具有重要地位。因此,苏铁群落区系组成的研究,对于古气候、古地理、种子植物起源演化以及现代植被类型的形成过程等研究都具有重要的意义。
1 研究区概况
攀枝花苏铁国家级自然保护区位于金沙江北岸的巴关河西坡及格里坪后山,东与巴关河及观音岩至胡家岩小路为界,与乌盐公路隔河相望;南与攀钢石灰矿与石华公路相邻。地理坐标:东经101°32'15″~101°35'46″,北纬 26°36'31″~26°38'24″。垂直分布幅度较宽,在海拔1 170 m~2 000 m均有分布,以1 500 m~1 700 m地段分布最为集中。分布上限接近山顶,下限在巴关河西坡可达河谷边缘。在格里坪后山,干热河谷气候影响显著,沿江一带发育为干热河谷灌丛或草坡,上限即接苏铁群落,保护区面积为1 358.3 hm2,区内约有24万株攀枝花苏铁。
区域地处金沙江下游河谷地带,河谷深切,地势低凹,呈封闭状,焚风作用显著,气候干燥炎热,使这里成为了南亚热带气候区的一块“飞地”。海拔1 500 m以下属南亚热带半干旱气候,该区全年无冬季,夏季长达190 d以上,年均温21℃,≥10℃积温7 500℃,最热月均温27.6℃,极高温41℃,最冷月均温13℃,极低温0℃;年降雨量800 mm,雨量集中于6月~9月,占全年降雨量的92.0%;年蒸发量为年降雨量的3倍以上,3月~5月最干燥,相对湿度低于40.0%,干燥度1.5~3.5。海拔1 500 m以上,属中亚热带半湿润气候,气候干热特征不太明显,年均温15℃ ~19℃最冷月均温6℃ ~10℃,干燥度1.0~1.49,≥10℃积温5 100℃ ~6 000℃,年降雨量800 mm~1 000 mm。
2 材料与方法
在保护区内有攀枝花苏铁生长的地段,根据坡向与植被外观的不同,分为阳坡矮灌丛,半阳坡高灌丛,阴坡稀树灌丛。对设置的样方中进行全员调查,并辅以多条样线调查,记录所见物种。并参考前人已有调查资料[8,9],予以补充完善;由此编订攀枝花苏铁群落植物名录。在此需要特别说明的是,苏铁群落的植物名录并非攀枝花苏铁自然保护区植物名录,那些栽培种、引入种和并不在苏铁分布地段出现的物种均被剔除;最后,统计出攀枝花苏铁群落中种子植物共有82科253属318种,并根据吴征镒对中国种子植物区系分布类型的划分进行区系分析[10]。
3 结果与分析
3.1 区系分布类型
3.1.1 科的分布类型
除世界分布的19个科外,泛热带科主要有防己科Menrispermaceae、锦葵科Malvaceae、爵床科Acan-thaceae、壳斗科 Fagaceae、萝摩科 Asclepiadaceae、马鞭草科Verbenaceae、木棉科Bombacaceae、西番莲科Passifloraceae、莲叶桐科 Hernandiaceae、番荔枝科Annonaceae、兰科Orchidaceae等44个科。热带亚洲和热带美洲间断分布有椴树科Tiliaceae和仙人掌科Cactaceae;旧世界热带分布有紫金牛科Myrsinaceae和芭蕉科Musaceae;热带亚洲和热带大洋州间断分布有百部科Stemonaceae与苏铁科Cycadaceae;热带亚洲(印度-马来西亚)分布为姜科Zingiberaceae。
温带成分中北温带分布有松科Pinaceae、伞形科Umbelliferae、报春花科Primulaceae、紫草科Boraginaceae、忍冬科 Caprifoliaceae、蓼科 Polygonaceae、毛茛科Ranunculaceae等10个科。另外,东亚及北美间断分布有小檗科Berberidaceae,旧世界温带分布有川续断科Dipsacaceae。

表1 攀枝花苏铁群落种子植物区系分布类型Table 1 The areal-types of spermatophytic families and genera of Cycas panzhihuaensis assemblage
3.1.2 属的分布类型
攀枝花苏铁群落区系组成中热带成分共计有151个属,比例为65.65%(世界分类型不计算在内)。其中泛热带分布类型主要有算盘子属Glochidion、麻疯树属Jatropha、美登木属Maytenus、车桑子属Dodonaea、山黄麻属Trema、刺蒴麻属Triumfetta、美冠兰属Eulophia、鱼黄草属Merremia、假酸浆属Nicandra、假杜鹃属Barleria、羊蹄甲属Bauhinia、薯蓣属Dioscorea、猪屎豆属Crotalaria、西番莲属Passiflora、榄仁属Terminalia等73个属。热带亚洲和热带美洲间断分布叶子花属Bougainviliea、雀梅藤属Sageretia、过江藤属Phyla、仙人掌属Opuntia、番石榴属Psidium、胜红蓟属Ageratum和羽芒菊属Tridax共12个属。旧世界热带分布类型有百蕊草属Thesium、千斤拔属Moghania、扁担杆属Grewia、吊灯花属Ceropegia、独脚金属Striga、飞蛾藤属Porana、香茶菜属Isodon、鸡脚参属Orthosiphon、石梓属Gmelina、细柄草属Capillipedium、拟金茅属Eulaliopsis等共17个属。热带亚洲和热带大洋州间断分布有苏铁属Cycas、樟属Cinnamomum、蝙蝠草属Christia、香椿属Toona、黑构叶属Leptopus、崖爬藤属Tetrastigma、茅瓜属Solena、荛花属Wikstroemia、野牡丹属Melastoma、百部属Stemona、山菅属Dianella共 11属。热带亚洲和热带非洲间断分布类型有凤凰木属Delonix、水麻属Debregeasia、木棉属Gossampinus、牛角瓜属Calotropis、南山藤属Dregea、青藤属Illigera、筒轴茅属Rottboellia、香茅属Cymbopogon、菅属Themeda、尾稃草属Urochloa、镰稃草属Harpachne等共25属。热带亚洲(印度-马来西亚)分布类型有青冈属Cyclobalanopsis、地黄连属Munronia、来江藤属Brandisia、杜根藤属Calophanoides、地皮消属Pararuellia、姜黄属Curcuma等13个组成。
温带成分共计56个属,所占比例合计24.35%(世界分类型不计算在内)。其中以北温带分布类型为最多,主要有松属Pinus、栎属Quercus、桑属Morus、荨麻属Urtica、翠雀属Delphinium、小檗属Berberis、漆树属Toxicodendron、白蜡属Fraxinus、琉璃草属Cynoglossum、蒿(艾)属Artemisia、天南星属Arisaema、黄精属Polygonatum、火烧兰属Epipactis等26个属。东亚及北美间断分布有栲属Castanopsis、石楠属Photinia、山蚂蟥属Desmodium、胡枝子属Lespedeza和蛇葡萄属Ampelopsis等12个属。旧世界温带分布类型有荞麦属Fagopyrum、火棘属Pyracantha、铜钱树属Paliurus、女贞属Ligustrum、香薷属Elsholtzia、川续断属Dipsacus、绵枣儿属Scilla、天名精属Carpesium共13属。温带亚洲分布有瓦松属Orostachys、杭子梢属Campylotropis和狼毒属Stellera共3个属。地中海区、西亚至中亚分布有沙针属Osyris和旱茅属Eremopogon。
另外,东亚分布类型和中国特有分布类型则属于华夏植物区系(Cathaysia Flora)成分。华夏植物区系是指三叠纪以来,在华南地台及其毗邻地区发展起来的被子植物区系[11]。华夏区系成分共计23个属,所占比例18.77%;尤以东亚分布类型为多,主要有油杉属Keteleeria、石莲属Sinocrassula、小石积属Osteomeles、九子母属Dobinea、阴行草属Siphonostegia、珊瑚苣苔属Corallodiscus、帚菊属Pertya、梧桐属Firmiana、象牙参属Roscoea、野丁香属Leptodermis、棕榈属Trachycarpus、舌喙兰属Hemipilia等19个属。中国特有属共4属,分别为翅茎草属Pterygiella、香果树属Emmenopterys、地涌金莲属Musella和栌菊木属Nouelia。
3.2 优势科与单属科统计
从表2可见,苏铁群落中优势科很少,含有20个以上属的科只有菊科和禾本科;含有10属以上的科仅有豆科和百合科。而单属科有39科,占总科数的47.56%;单属科中绝大多数也是单种科,有苏铁科、檀香科、马兜铃科、番荔枝科、木棉科、莲叶桐科、西番莲科、仙人掌科等33科,分别占到总科数的40.24%,占单属科总数的84.62%。单种科和单属科反映了植物科进化过程中相反的方向,一个是新产生的科,其属种尚未分化,尚没有新衍生的属种;另一个是演化终极的科,属种已经大量消亡,现存的是残遗种类。对单种科和单属科的分析可反映出一地区植物进化的历史和现状。由于苏铁群落区系中的单属科在系统分类上多处于原始类群,因此,它的单属科更多表现出一种古老性和残遗性现象。
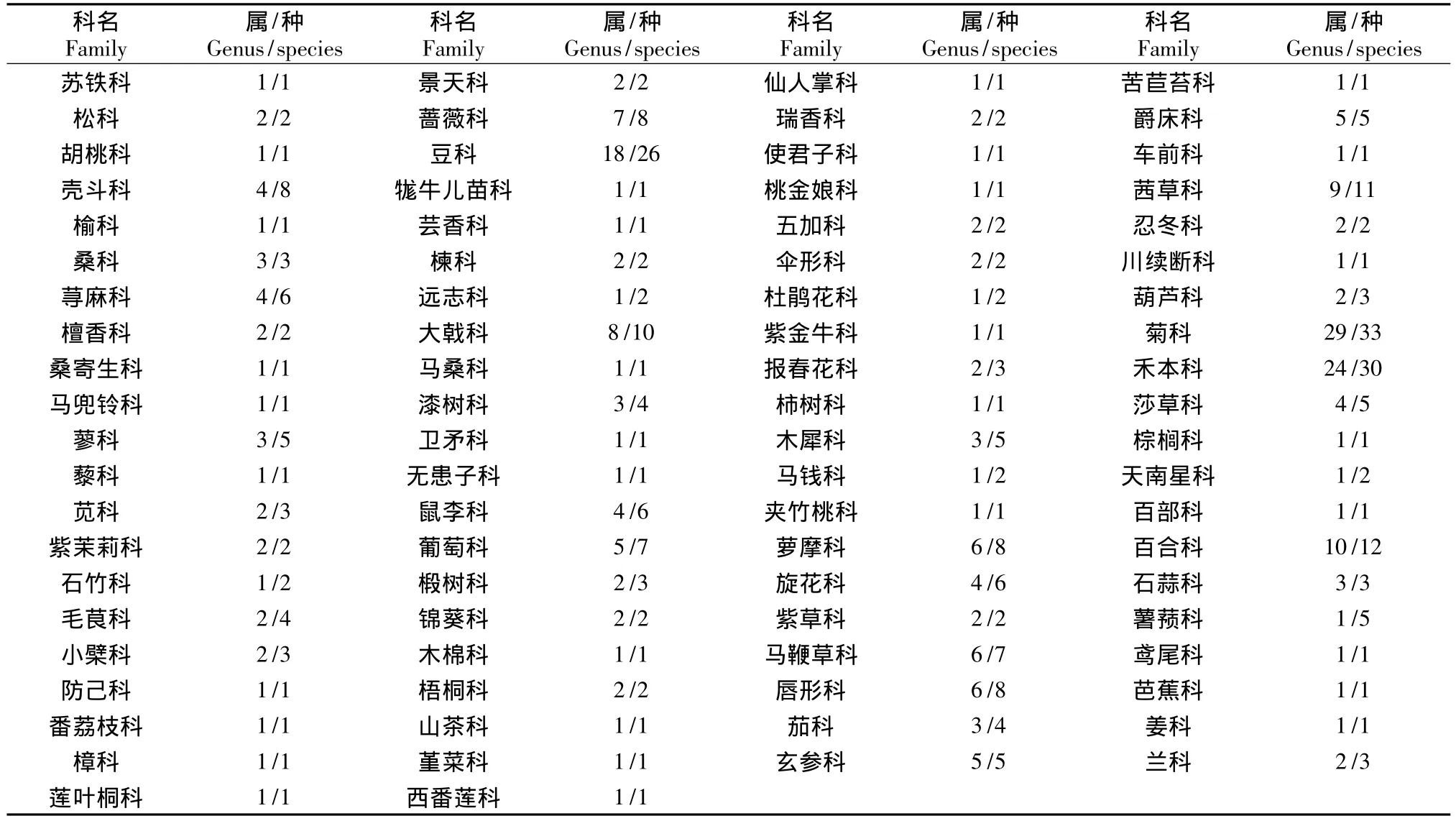
表2 各科所含属数和物种数Table 2 Statistics of genera and species of each family in Cycas panzhihuaensis assemblage
3.3 单种属与寡种属统计
属与种的比值大小在一定程度上反映出以属为单位的种系在历史发展过程中的长短,另一方面还存在着面积与属种的比值等因素。攀枝花苏铁区系中单种属达209个,占总属数的82.61%;其中大部分是中国特有种,部分为干热河谷特有种,反映出区系强烈的地带性特征,充分显示了区系的古老性和特殊性。寡种属(2种~3种)有40个,占总属数的15.81%;包含4个种的属仅有铁线莲和羊蹄甲属,所含种数最多的属是栎属和薯蓣属,它们属下也都只有5个种。可见,区系中以单种属和寡种属占有绝对优势,而无明显的优势属。
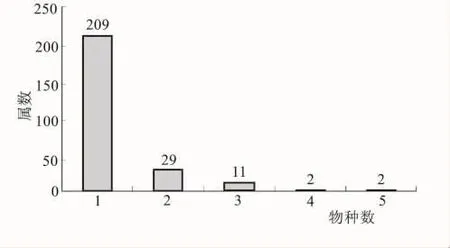
图1 属级大小与所含物种数统计Fig.1 Statistics of the size of the seed plants genus
3.4 特有种与标志种
标志植物是广义的特征植物,用以标志或表征一个自然区域的近代生境和相对较近代的区系历史及其地理分布特征[12]。植物区系的标志植物在种级可称标志种(或特征种),特有种因为它们的分布区都较窄,适应特有的地理环境而一般都是比较好的标志种。苏铁群落中共有干热河谷标志种93种,占到群落总种数的29.25%。其中攀枝花苏铁、草坡豆腐柴Premna steppicola、地涌金莲Musella lasiocarpa、金沙翠雀花Delphinium majus、栌菊木Nouelia insignis、金沙荆Vitex duclouxii、云南百部Stemona mairei、金江南星Arisaema jinshajiangense和金沙尾稃草Urochloa jinshanicola为金沙江干热河谷特有种[13],占到金沙江干热河谷特有种总数的28.13%。另外,非特有的标志种有81种,如假苜蓿Crotalaria medicaginea、尾稃草Urochloa reptaus、栎叶枇杷Eriobotrya prinoides、云南羊蹄甲Bauhinia yunnanensis、云南山蚂蝗Desmodium yunnanense、小檗美登木Maytenus berberoides、车桑子Dodonaea viscosa、须弥茜树(丽江山石楠)Himalrandia lichiangensis、余甘子Phyllanthus emblica、假杜鹃Barleria cristata、大独脚金Striga masuria、西南吊兰Chlorophytum nepalense、龙棕Trachycarpus nana、镰稃草Harpachne harpachnoides等。
另外,珍稀濒危植物除攀枝花苏铁外,还有云南梧桐Firmiana major、栌菊木和龙棕。其中云南梧桐为我国特有珍稀树种,曾一度被认为几乎野生灭绝[14]。世界保护联盟(IUCN)公布的1997年度世界受威胁的树木名录中,将云南梧桐列为野外绝灭的3种中国特有植物之一。而后又在攀枝花等干热河谷地带被相继发现有野外种群分布[15],其珍稀濒危程度不亚于苏铁类植物。
4 结论与讨论
4.1 孑遗植物区系组成的古老性与特有性
苏铁群落区系以热带成分为主,泛热带成分尤多,表明攀枝花苏铁群落具有强烈的热带性质和起源的古老性,也反映出它与亚洲的热带分布区类型存在较多的联系。攀枝花苏铁是古老裸子植物的残遗或后裔,起源于康滇古陆;栌菊木是菊科木本植物的原始种,也是干热河谷残遗的古老成分[6]。地区特有种地涌金莲是金沙江河谷特有的单型属种,与热带非洲至热带亚洲分布的象腿蕉Ensete十分接近;另外,如车桑子、铁仔Myrsine africana、清香木Pistacia weinmannifolia、沙针Osyris wightiana等常见种也是古地中海的残遗分子。
温带成分中硬叶常绿栎类树种是古地中海沿岸干旱生境硬叶常绿植物的后裔,其常绿种类在中国-喜马拉雅和地中海区得到充分发展;而林分中铁橡栎Quercus cocciferoides与产地中海东部的灌木栎Q.coccifera有形态特征上具有相似之处。这些温带成分反映了攀枝花苏铁群落的植物地理成分与古地中海区有着密切的联系,其起源和发生背景与古地中海有关[9]。温带成分的侵入主要得益于第四纪冰期活动,那时这里成为了第三纪以来古地中海沿岸古老植物及其后裔的避难所,众多类群在此得以残存、演化发展至今,共同造就了现今区系成分的古老性与特有性。
4.2 干热河谷性质的典型性与代表性
苏铁群落区系干热河谷性质明显,干热河谷标志种所占比例接近三分之一。一些物种为适应干热河谷的特殊生境已分化出特化类型;如芭蕉科高大草本植物在这一地区却出现了矮小草本种类型—地涌金莲[16]。龙棕是棕榈属中为数不多的无主茎的物种;九子母属是漆树科中唯一生长型为亚灌木草本的属,表明这也是从乔木分化出来的过渡类型。这些植株矮化的特化现象暗示着诸多类群在这个“分化中心”地区由木本向草本过渡;这就印证了西南地区是中国植物区系的分化中心,同样也是苏铁类植物的分化中心。
4.3 与古地理、古植物区系演化关系
攀枝花苏铁群落分布区位于中国亚热带西部半湿润常绿阔叶林区,植物区系属中国-喜马拉雅区的东缘,与中国-日本植物区相衔接,是两大植物区系的交汇点。如中国喜马拉雅分布的九子母属仅有2个种,苏铁群落中的羊角天麻D.delavayi与东喜马拉雅的贡山九子母D.vulgaris呈间断分布;又如红椿Toona ciliata、云南黄杞Engelhardtia spicata、榕树Ficussp.、刺蒴麻Triumfettasp.、榄仁Terminaliasp.等一些种类与喜马拉雅植物区系也存在一定联系。这都印证了云南西北部至四川金沙江河谷一带可能是中国-喜马拉雅和中国-日本两个东亚分布变型的北界,同时也可能就是古地中海的真正边界的论断[16]。甚至也有推测:“横断山脉包括川西这一块地区,可能是东亚许多植物分类分化的摇篮,还可能是某些植物分类群的始生中心,尤其是西与喜马拉雅,东至台湾显示出密切的亲缘”[17]。
自中生代以来,康滇古陆较稳定的古地理环境为苏铁植物发生发展提供了良好条件,而中生代三叠纪正是种子蕨开始衰退和苏铁植物开始大发展的时期,康滇古陆及其邻近山地是苏铁类植物化石群的主要产地。晚三叠世考依卜期在川西南金沙江大弯曲地区出现了我国最早的重要苏铁植物群,也是世界上最早的苏铁类植物群之一,即攀枝花市的宝鼎地区大荞地组[18,19]与盐边红坭组植物群[20]。该化石植物群中苏铁类植物占到全部植物种类的一半以上;现今的攀枝花苏铁便是其中塔状铁雄果Androstrobus pagodiformis和硬叶焦羽叶Nilssonia sclerophylla的直接后裔[21]。另外,攀枝花市晚三叠世植物群的种子蕨植物化石也相当丰富[22]。植物的发生、发展与迁移是与整个植物群密切相关;苏铁植物的发生与种子蕨有着直接的亲缘关系,它们是古生代类型的延续,充分显示了植物类群的古老性。甚至可以推测这一地区很可能就是世界苏铁植物重要的起源与发展中心[9]。
[1] 朱家柟,杜贤铭.中国始苏铁(新属、种)Primocycas chinensisgen.et sp.nov.在我国早二叠世的发现及其意义[J].植物学报,1981,23(5):401 ~404.
[2] 陈家瑞.中国苏铁[J].植物杂志,1996,(2):3 ~6.
[3] 周林,杨思源,傅立国,等.在四川发现两种新苏铁[J].植物分类学报,1981,19(3):335 ~338.
[4] 孙芝和.四川省宁南发现攀枝花苏铁[J].生命世界,1984,3:13.
[5] 何永华,李朝銮.攀枝花苏铁种群生态地理分布、分布格局及采挖历史的研究[J].植物生态学报,1999,23(1):23 ~30.
[6] 吴征镒.论中国植物区系的分区问题[J].云南植物研究,1979,1(1):1 ~22.
[7] Li H L.Floristic relationships between eastern Asia and eastern North America[J].Transactions of the American Philosophical Society(New Series),1952,42:371 ~429.
[8] 周立江,管中天.金沙江河谷苏铁天然植物群落的研究[J].云南植物研究,1985,7(2):153 ~168.
[9] 管中天.中国苏铁植物[M].成都:四川科学技术出版社,1996.
[10] 吴征镒,路安民,汤彦承,等.中国被子植物科属综论[M].北京:科学出版社,2003.
[11] 张宏达.华夏植物区系的起源与发展[J].中山大学学报,1980,19(1):89 ~98.
[12] 金振洲,欧晓昆,区普定,等.金沙江干热河谷种子植物区系特征的初探[J].云南植物研究,1994,16(1):1 ~16.
[13] 金振洲,杨永平,陶国达.华西南干热河谷种子植物区系的特征、性质和起源[J].云南植物研究,1995,17(2):129 ~143.
[14] 傅立国,等.《中国植物红皮书》(第一卷)[M].北京:科学出版社,1992.
[15] 王大绍.攀枝花发现云南梧桐天然林[J].植物杂志,2001:4~5.
[16] 吴征镒,王荷生.中国自然地理—植物地理(上)[M].北京:科学出版社,1983.
[17] 管中天.四川松杉植物地理[M].成都:四川科学出版社,1982.
[18] 李佩娟,曹正尧,吴舜卿.《云南中生代植物》云南中生代化石(上册)[M].北京:科学出版社,1976.
[19] 杨贤河.中生代植物界.《西南地区古生物图册》,四川分册(二).地质出版社,1978.
[20] 陈晔,段淑英,张玉成.四川盐边晚三叠世植物新种[J].植物学报,1979,21:1 ~2.
[21] 胡雨帆.攀枝花三叠纪苏铁植物化石居群的特征及其生态学意义[A].中国古生物学会第21届学术年会论文摘要集[C].2001.
[22] 徐仁,等.中国晚三叠世宝鼎植物群.北京:科学出版社,1979.
