坡形因素对麻疯树人工林个体与群体生物量配置的影响
2011-06-26何亚平费世民蔡小虎王乐辉陈秀明蒋建强雷彻虹
何亚平,费世民**,蔡小虎,王 鹏,王乐辉,陈秀明,蒋建强,雷彻虹
1.四川省林业科学研究院,四川 成都 610081;2.长江造林局攀枝花分局,四川 攀枝花 617000;3.攀枝花市林业局,四川 攀枝花 617000
坡形(slope forms)是自然立地的一类重要特征,反应了立地的水平面上曲线特征。在三维空间中坡形是曲面,在二维空间中坡形是曲线;坡形分为直线性坡形和曲线性坡形两类,曲线性坡形又分凸形、凹形和S形坡等;坡形变化复杂的可称为复合形坡。凸形坡(convex slope)表示坡面呈一上凸的曲线,表明山体浑圆,坡上部平缓,下部较陡;凹形坡(concave slope)表示坡面呈一下凹的曲线,表明山体较陡,尤其是上部更为陡峭;所谓复合形坡(composite slope)表示坡形有时呈拉长了的“S”形,即坡上部浑圆而上凸,下部陡而下凹等。坡形反应了自然立地下物质配置规律,是物质迁移、水分运动与植被动态长期相互作用的产物。坡形也是不同立地条件的组合体,具有环境特性。不同类坡形之间具有小尺度物质运输上的相互联系,并存在与之相对应的生物特性。坡形条件下生物活动规律研究少,还不清楚坡形对生物活动的影响或生物对不同坡形的适应性。
坡形也是人工栽培基地的基本特征。麻疯树生物柴油原料工业人工林定向培育离不开规模化基地建设与高效栽培。在相同的栽培措施下,坡形不同,麻疯树大小不同,其产量也不同。麻疯树个体和群体水平上,都对坡形产生了适应性,塑造基地个体、群体或景观斑块异质性。坡形与动植物的异质性复合体,也是微观尺度上生态系统类型划分的基本依据。基地尺度上定向培育与高产稳产需要最大化每一单株、群体的生长与生殖活动,或提高劣势坡形条件下的生长水平与资源利用效率,优化基地景观结构与经营活动,进而从整体上提高种子生产能力。不同坡形条件下麻疯树生长量与种子生产能力研究利于认识坡形这一工业人工林基地中普遍存在的立地特征与生物效应现象,并为高产稳产定向培育提供基本的生物生态学信息。
相关研究也有所报道,如卢建国等(2006)研究了油蒿种群生物量对水分异质性的适应;Li等(2005)研究灌木Artemisia halodendron生殖配置的生境异质性;郭永清等(2010)、莫丽芬等(2010)研究麻疯树幼林生物量、生产力与种子产量。罗建勋等(2007)研究了麻疯树种子产量与坡位、坡形、土壤厚度的相关性。这些研究都未涉及到麻疯树基地坡形异质性对生物量、生产力、生殖配置与种子产量的影响或效应问题,限制了经营技术认识。
本文研究了金沙江干热河谷攀枝花段一个麻疯树人工林不同坡形条件下个体与群体生物量配置,认识坡形对工业原料林的生物生态学效应,为经营模式与坡形立地优化配置研究提供理论依据。选择2006年建设的马店河基地,测定斜坡、平地、缓坡、山坳与凸坡几种条件下个体与群体生物量、生产力与种子产量,并分析其器官配置状况。
1 材料与方法
1.1 试验地概况
生物量测定林地位于攀枝花市仁和区大龙潭乡马店河,海拔 940 m,经度为 101°51'601″,维度为26°28'132″。试验区域为西南山地金沙江干热河谷攀枝花段,处于元谋干热河谷向雅砻江流域干暖河谷过渡地带,在宽谷江边为稀树灌木草丛,峡谷地带为灌木林。该地处金沙江干流中山丘陵区,坡度在5度~45度之间,坡位为中坡,坡向南,为第二台,土壤为山地红壤,粘粒紧实,坡积土,母岩为砂岩。马店河人工林自然植被为稀树灌木草丛,灌木树种有台湾相思、蜜柚子、麻疯树,造林后以麻疯树为主体;草本植物主要是双花草、扭黄茅,在积水湿地上有莎草与苔草属植物。
1.2 林地概况
马店河麻疯树人工林为2005年栽植,栽植坑大小为40 cm×40 cm,栽植当年施底肥,2006年、2007年追施复合肥,2008年施配方肥(主要是基地下部,进行配方专用肥试验),2009年施配方肥(全基地施肥,分夏、冬两季施予)。抚育管理措施是除草做防火带,人为活动是放牧,存在明显的土壤干扰,形成斑块状的积水地,并使得树体存在明显的擦痕,存在一定数量的断枝、枯死枝。因放牧导致的林地积水,该部分林分个体在水干后因土壤板结而缺乏氧气进而根系生长受限,导致营养合成不足几乎无结实现象或存在极少量结实。
1.3 数据测定
选择林地代表性立地5个,样地基本特征如表1所示,分别选择10株~15株测定来作为生物生产量测定样品。2008年春季4月份标记个体,并把株高划分为7级做标记登记枝径(下部第一级为基径),4月~5月份统计雌、雄花比例,7月~8月份统计果实数量,1月份落叶后测定冠幅、树高、再次登记枝径、收获全株,并依据叶痕、节龄痕来分别统计叶片总数、不同年龄枝条数量,分别称每个年龄段上的鲜重。选择每个立地选择干叶片30片,室内测定单叶干重。收割时收集根系鲜样品、3 a生枝鲜样品、1 a生枝条鲜样品来获取水分含量并计算干枝根系的干重。去掉破坏的个体后,共计获得45生物量测定个体与39个生产力测定个体。生殖器官生物量主要为凋落的雄花、果实、果序几部分,花通过原有的单花重量、性比(果实数量可代表雌花数量)、单果重量来估算干重,果实、果序重要通过果实数量 与成熟果序质量之间的模型来计算。

表1 马店河麻疯树基地不同坡形样地的基本特征Table 1 The general feature of plots with different slope forms in Madianhe J.curcas base
1.4 生产力估算方法
木本植物生产力测定难度在于区分积累性维持器官部分的比重,新生部分可直接区分并测算产量,如叶片、生殖器官与新梢。麻疯树枝干部分年龄痕较为明显,也可用年龄分段平均粗略估算当年生产量。麻疯树生产力估算方法采用两种方法来计算:
(1)直径增量
通过生长季节开始时直径与生长季结束时直径构成的同心圆来计算圆环部分生产量估算积累器官的生产量,而临时性器官则直接累加产量即可。
(2)年龄分段平均
干枝根部分不同年龄表示生长物质积累了多少年,则单年实际生长量可简单计算为每年的平均值。年龄分段平均估测生产力的方法在毛竹生产力研究中采用(何亚平等,2007),该物种克隆个体可作为不同年龄的枝条。但每年生长量不同,各器官积累也可能不同,只能为粗略估算。
1.5 数据计算与分析
计算不同坡形条件下个体生物量配置与群体生物量密度,绘制柱状图。生殖配置分别有两类,一类是生物量计算出的生殖配置,一类是生产力计算出的生殖配置(成为生殖配置1和生殖配置2)。群体生物量密度用实际密度与样木来计算。
2 结果与分析
2.1 不同坡形条件下的结构特征
不同坡形条件下个体地径、树高、冠直径、冠幅、枝下高不同(如表2所示)。斜坡地直径最大,缓坡地、山坳、凸坡次之,平地直径最小,呈现斜坡地>缓坡地>山坳>凸坡>平地的模式。冠直径、冠幅参数都表现出斜坡地最大,缓坡地、山坳次之,凸坡与平地相似。树高斜坡地最大,缓坡地与山坳相似,而凸坡与平地相似。平地枝下高较大,凸坡、山坳次之,斜坡地、缓坡地较小。个体形态参数表明,斜坡地地径、树高、冠幅参数较大,而枝下高较小,缓坡地、山坳依次降低,凸坡、平地最低(树高参数在山坳与缓地坡地稍微相似)。
林分密度在不同坡形条件下不同,斜坡地密度最大(240株·0.067 hm-2),平地最低(140株·0.067 hm-2),凸坡(213 株·0.067 hm-2)、山坳(180株·0.067 hm-2)、缓坡地(153株·0.067 hm-2)次之。造林设计密度相同(107株·0.067 hm-2),但造林时因坡度(测算为垂直投影)、石块分布等因素会造成实际密度偏差。

表2 马店河麻疯树基地不同坡形条件下麻疯树个体结构特征Table 2 The individual phenotypic feature of plots with different slope forms in Madianhe J.curcas base
2.2不同坡形条件下个体生物量的绝对配置
不同坡形条件下麻疯树单木生物量绝对配置不同(如表3所示)。叶片生物量为斜坡地>缓坡地>山坳>平地≈凸坡,干枝生物量为斜坡地>缓坡地>山坳>凸坡>平地,根系生物量为斜坡地>缓坡地>山坳>凸坡>平地,总体生物量为斜坡地>缓坡地>山坳>凸坡>平地。生殖相关参数在坡形间差异明显不同,生殖生物量为斜坡地 >山坳>缓坡地>凸坡>平地,果实数量为山坳≈斜坡地>缓坡地>凸坡>平地,种子产量为斜坡地>山坳>缓坡地>凸坡>平地。显然,生殖生物量、果实数量与种子产量大小顺序与营养生物量不一致,山坳比缓坡地要高一些;果实数量山坳与斜坡地相似。
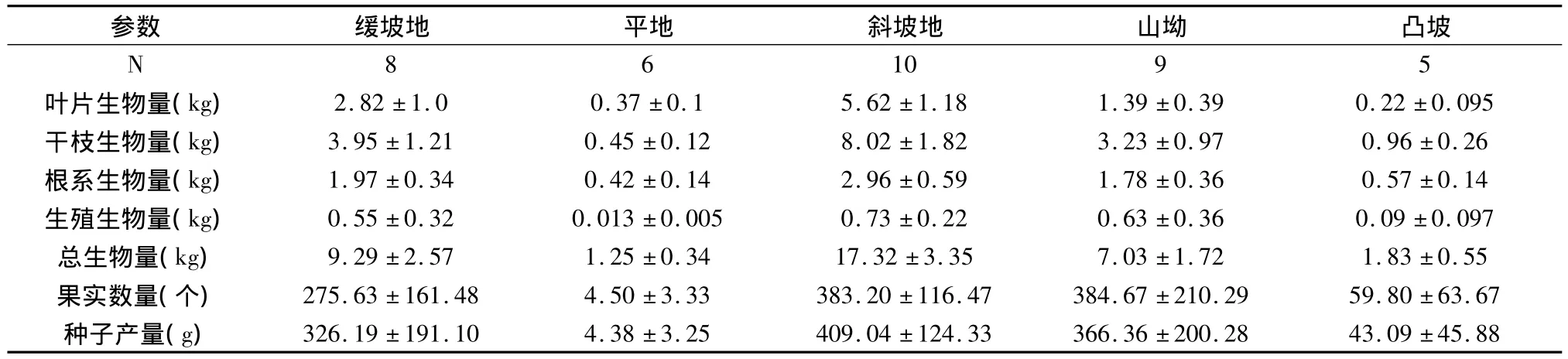
表3 不同坡形条件下麻疯树单木生物量绝对配置Table 3 The absolute allocation of individual biomass in plots with different slope forms in Madianhe J.curcas base
统计分析表明,叶片生物量斜坡地最高,缓坡地次之,山坳较低,而平地、凸坡最低,呈现四个明显差异的高度等级;干枝生物量与叶片生物量显著性差异等级划分类似。根系生物量在斜坡地最高,缓坡地与山坳次之,平地与凸坡较低,呈现3个明显差异等级。生殖生物量缓坡地、斜坡地、山坳最高,平地与凸坡较低,呈现两个显著性差异等级。斜坡地最高,缓坡地与山坳次之,平地与凸坡最低,呈现3个明显差异等级。果实数量缓坡地、斜坡地与山坳最高,而平地与凸坡最低,呈现两个显著的等级。显著性分析柱状图如图1所示,生殖生物量、果实数量与种子产量柱状图等级相似,叶片与干枝生物量相似,根系生物量与总生物量相似。
2.3 不同坡形下生物量的相对配置
坡形不同,生物量相对配置不同,坡形之间差异因器官而异(如表4、图2所示)。叶片配置:斜坡地≈缓坡地≈平地>山坳>凸坡;干枝配置:凸坡>山坳≈斜坡地≈缓坡地>平地;根系配置:凸坡≈平地>山坳≈缓坡地>斜坡地;生殖配置:山坳>缓坡地≈斜坡地≈凸坡>平地。叶片配置斜坡地、缓坡地与平地相对较高,干枝配置凸坡、山坳相对较高,根系配置凸坡、平地相对较高,而生殖配置则是山坳、缓坡地、斜坡地相对较高。

表4 不同坡形条件下麻疯树单木生物量相对配置Table 4 The relative allocation of individual biomass in plots with different slope forms in Madianhe Jatropha curcas base
通过生产力计算叶片配置与生殖配置分别称之为叶片配置1与2,生殖配置1与2。叶片生殖配置1:平地≈斜坡地≈缓坡地>山坳>凸坡;叶片生殖配置2:缓坡地≈平地≈斜坡地>山坳>凸坡;生殖配置1:山坳>缓坡地≈凸坡≈斜坡地>平地;生殖配置2:山坳>缓坡地≈凸坡≈斜坡地>平地。
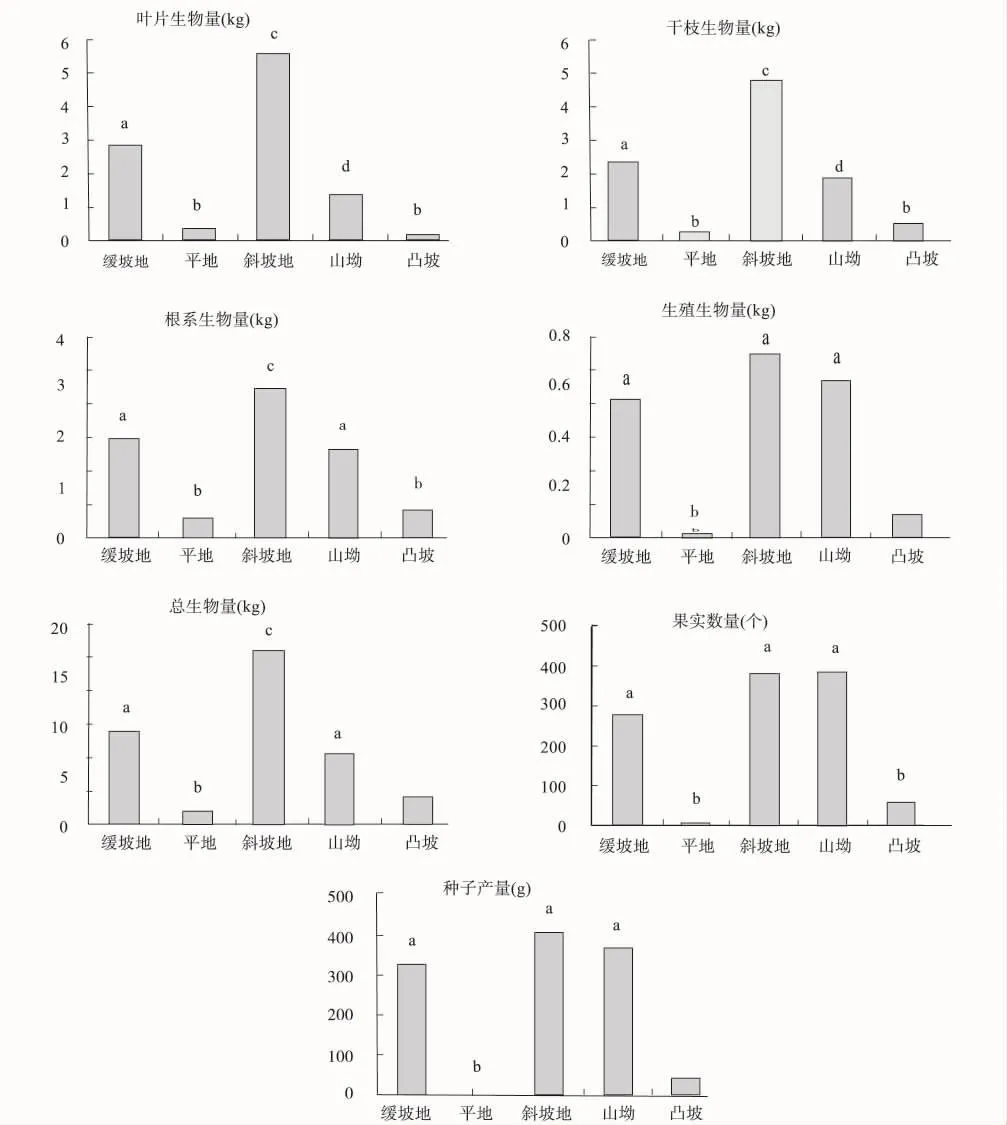
图1 不同坡形条件下麻疯树单株生物量绝对配置与种子产量Fig.1 The absolute allocation of individual biomass of Jatraphas curcas and its seed yield under the coanditions of different slope forms
2.4 不同坡形群体生物量与生产力
不同坡形群体生物量及其组成如表5所示。总体生物量斜坡地最大,缓坡地、山坳相对较低,凸坡与平地最低,坡形间差异极为明显(df=4,f=15.632,p=0.000)。叶片生物量斜坡地最高,缓坡地次之,山坳较低,平地与凸坡最低,叶片生物量在坡形间呈现明显差异(df=4,f=89.037,p=0.000)。干枝生物量斜坡地最高,缓坡地与山坳次之,凸坡较低,而平地最低,坡形间干枝生物量差异明显(df=4,f=68.539,p=0.000)。根系生物量斜坡地最高,山坳与缓坡地次之,凸坡较低,而平地最低,坡形间根系生物量差异明显(df=4,f=71.52,p=0.000)。生殖生物量斜坡地最高,山坳与缓坡地次之,凸坡与平地最低,坡形间生殖生物量差异明显(df=4,f=90.654,p=0.000)。马店河4 a生人工林不同坡形群体生物量浮动在2.63 t·hm-2~62.34 t·hm-2之间,其中干枝生物量最大,叶片次之,根系较低,而生殖生物量最低。
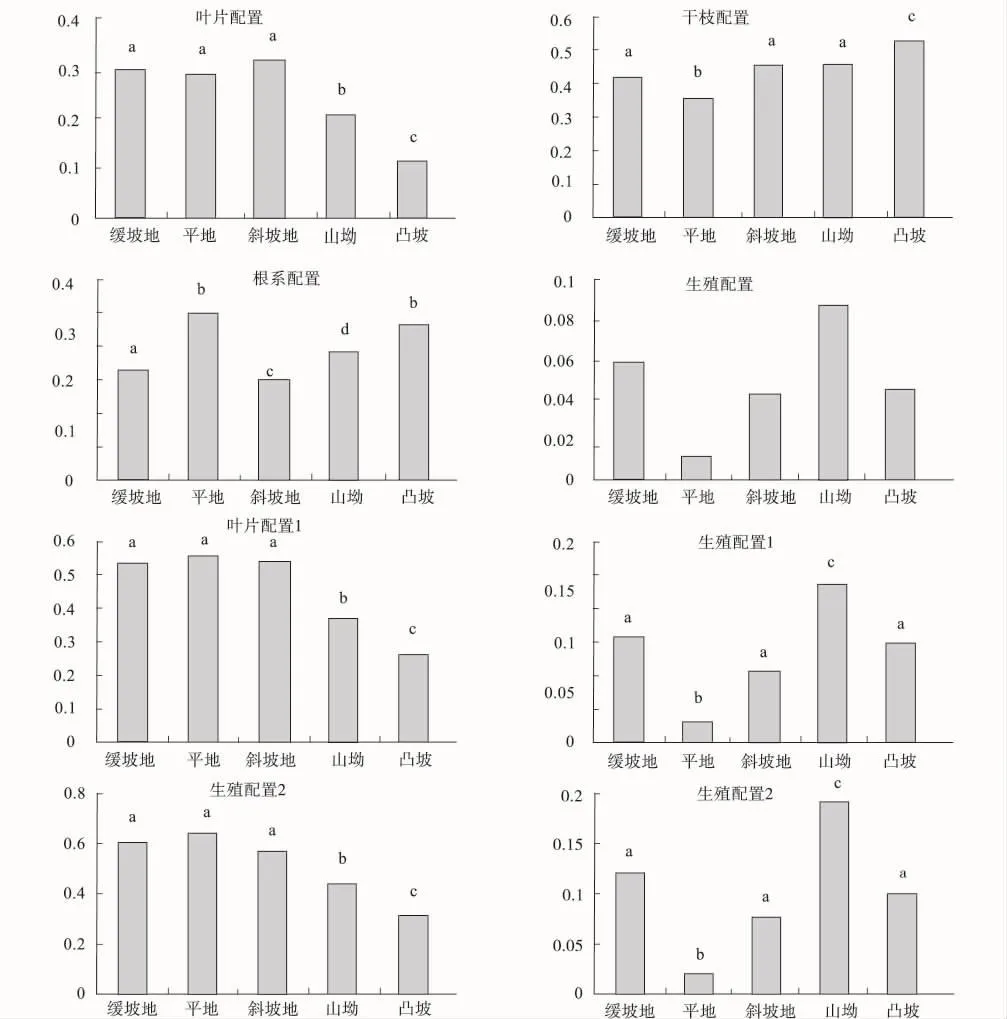
图2 不同坡形条件下麻疯树单株生物量的相对配置Fig.2 The relative allocation of individual J.curcas biomass in different slope-form plots in Madianhe base
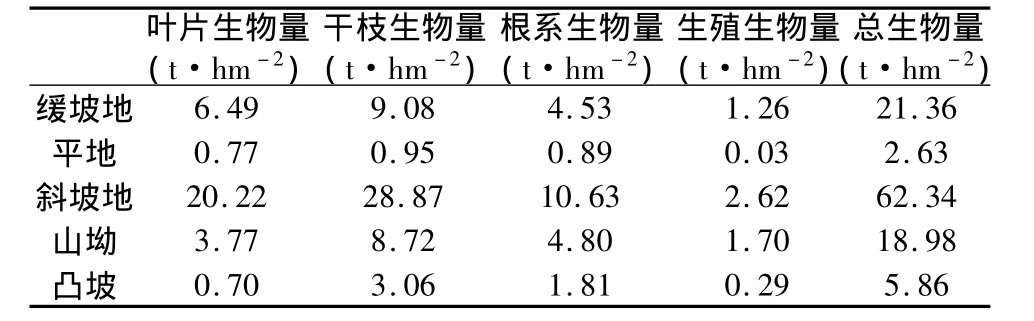
表5 不同坡形条件下麻疯树群体生物量Table 5 The population biomass under the conditions of different slope forms in Madianhe J.curcas base
坡形间群体生产力如表6所示。缓坡地生产力(年龄分段法)斜坡地最高,缓坡地与山坳次之,平地与凸坡最低,坡形间生产力差异极为明显(df=4,f=80.211,p=0.000)。增量法计算的群体生产力与年龄分段法计算的大小一致性相似,坡形间也存在极显著差异(df=4,f=79.184,p=0.000)。马店河4a生人工林生产力浮动在1.28 t·hm-2·a-1~37.15 t·hm-2·a-1之间。

表6 不同坡形群体生产力与种子产量Table 6 The population productivity and seed production under different slope forms
2.5 不同坡形群体种子产量
不同坡形群体种子产量如表6所示。斜坡地与山坳果实数量最多,分别为1 379.50×103个与1 038.6×103个(约130与100万个);缓坡地果实数量次之,为633.94×103个(约63万个),凸坡与平地果实数量相对较低,分别为191.36×103个与9.45×103个(约为10万个与1万个)。不同坡形果实产量差异极为明显,检验达到极显著水平(df=4,f=15.052,p=0.000)。
群体种子产量在坡形之间差异极为明显(df=4,f=15.978,p=0.000),斜坡地种子产量最大,每公顷达到1 472.5 kg;山坳种子产量次之,每hm2达到989.18 kg;缓坡地种子产量相对较低,每hm2为750.24 kg;凸坡与平地种子产量最低,分别为137.90 kg·hm-2与 9.21 kg·hm-2。马店河 4 a 人工林种子产量浮动在9.21 kg·hm-2~1 472.5 kg·hm-2。
3 讨论
麻疯树规模化基地内部景观结构并不是整齐划一的,能源林生长呈现异质性格局,这种异质性抛开种质遗传差异而单纯考虑生态因素,则是坡形主导的水文格局造成的。生物量测定表明,平地与凸坡生物量较低,个体较小,生产量较低;缓坡地生物量相对较高,个体较大,生产量提高,山坳与斜坡地生物量最高,个体最大,生产量最高。显然,基地产量内部呈现出异质性斑块格局,如斜坡地、缓坡地、小山坡较高,而平地、凸坡较低。这种坡形主导的产量异质性与营养生长有关,而实质上是坡形条件不同时资源丰沛性不同所致。坡形直接影响水文过程,可能对壤内流、蓄积、径流与蒸发影响不同,致使不同坡形条件下土壤可利用水分含量不同。不同水分含量直接导致营养生长量与积累量不同,形态参数、生物量与生产力不同,进而生殖产量不同。卢建国等(2006)研究毛乌素沙地土壤湿度异质性与油蒿种群空间异质性之间关系,表明油蒿生物量与土壤湿度呈现显著正相关关系,并且地形的斑块性→湿度斑块性→油蒿种群分布的斑块性→生物量的斑块性。坡形对麻疯树分布与生物量、生殖产量异质性的塑造作用与坡形异质性存在必然联系。试验证明,随着土壤水分含量降低,疯树的株高、生物量均呈现下降趋势(焦娟玉等,2010),对水分反应明显。
坡形导致的能源林生产异质性格局对基地高产稳产经营管理策略影响明显,可能驱动经营者针对不同坡形而采取相应的措施。但需要认清坡形条件下资源本质与规律便于提炼技术模块,集成创新麻疯树经营模式与技术体系。大气降水是规模化基地水分与物质输入的主要途径,肥力还有一个来源是岩石风化,但物质通过水文驱动在坡面发生迁移,在重力作用下从凸坡向下迁移,并沿途经过截留吸附与下渗,而在平地、缓坡地、凹坡地带形成水肥富集地带,而凸坡则水肥逐渐贫瘠。凸坡风力强劲,蒸发大而散失快,水分保蓄不足,干旱明显;平地凹坡积水明显,旱季雨涝而冬季土壤板结不透气。缓坡地、斜坡地与小山坡(大凹坡中间的小凸坡)疏水性良好,但蒸发也没有凸坡那么大,水分相对适宜。麻疯树基地非生物资源坡形差异也是权衡配置的结果,而坡形异质性是驱动权衡配置的必须条件。资源权衡配置而不是均质化配置是驱动异质性与维持生命活力的一种自然法则。单纯考虑工业人工林,则麻疯树基地坡形异质性而不是均质性驱动了景观斑块的异质化,不利于高产稳产;基地定向培育需要考虑坡形异质性的生物学效应,科学合理配置现有技术资源,从立地控制、结构控制与遗传控制上入手,适地适树适结构适模式并适效益,优化组合,最终促成种子产量形成的高产稳产。目前,有关坡形的生物生态学效应研究很少,限制了异质性下“地-树-结构-模式”技术模式效益思维的运用,这也是长期以来忽略了空间生态学而忽视生态学基本原理的结果。坡形对形态结构参数、生物量与器官配置、生产力,以及种子产量等影响还受到营养器官与生殖器官之间权衡,器官发育的异速生长定律的调节。坡形对生长与生殖产量的影响也是先通过根系限制来影响营养生长进而影响生殖生长(何亚平等,2010)。
个体及其器官生产量之间相关分析表明(如表7所示),生殖生物量与叶片、干枝、根系、总生物量、根系与生殖生物量相对配置、生殖配置1与2、果实数量、种子产量紧密相关,而与叶片配置、干枝配置无关;表明,生殖生物量直接影响单株果实、种子产量,与生殖、根系比例有关,而与叶片、干枝比例无关,但都与各器官生物量积累量有关。单纯考虑种子产量,则都与叶片、干枝比例无关,而与器官生物量、根系与生殖比例相关。单株各器官生物量储量都与种子产量、生殖生物量呈现正相关,但与叶片、干枝比例却无关,表明叶片比重、干枝比重可能不是促进生殖的关键因素,可能树体内部干枝与叶片生物量优化配置才是关键,但干枝与叶片总量越大越利于种子生产。
叶片生产利于新枝发育,而生殖器官发育在头年枝发育相关而呈现头年枝大小依赖性(一定上随着枝条大小增加而增强)(何亚平等,2008,2009),因而叶片生物量大则利于当年后续与来年生殖器官发育。叶片与生殖器官总体发育联系紧密。干枝直接孕育与承载生殖器官发育,决定了头茬与后续二、三茬生殖器官发育,与生殖生物量与种子生产联系紧密。叶片与干枝这一生殖器官直接联系器官在整体中的比重则并不能代表资源配置的所有,其比重与营养生长有关,并不能反应内部枝条数量与质量优化程度,或者现有测试条件下枝条未呈现优化配置而无关。显然,营养器官生物量与生殖生物量、种子产量之间呈现正线性关系,表明生殖与种子产量是大小依赖的,营养生物量提高是提高种子产量的基础条件,但营养生物量优化配置而降低消耗器官而增加储存器官,保持适宜的构件资源优化配置度,追求单株产量的高产与稳产。目前树体管理经验局限于果树,虽有参考价值,也是原理相同而运用不同。麻疯树喜光而营养枝发育受到个体、构件密度效应影响而效果不佳,局部修枝效果不明显(马店河路下、左上、左上上等)。试验研究中采用整体截干而枝条发育极好,培育优势壮枝,来优化营养枝生物量配置,这在岔河基地随处可见。但出现了枝条过壮需要增加密度来优化质量,来增加一级枝角度,培育来年果枝,并在壮枝发育呈现密度效应时,果实成熟后迅速优化结构,隔级修枝,在采穗圃穗条培育上“截干-修枝”基础上,提出了隔级修枝技术,这与园艺上与果树上树体管理技术截然不同。这一差异也是生殖生产特殊性与麻疯树树种生物生态学特性不同所致。这种追求生殖生物量与种子产量而优化营养器官配置的思路也是优化结构度而依度营养的基本点,是生殖调控的基本途径。
相关分析也表明,生殖生物量相对配置与生殖生物量紧密相关,而与叶片生物量、干枝生物量、根系生物量、总生物量,以及叶片配置、干枝配置、根系配置无关;生殖配置1与生殖配置2也是如此(如表7所示)。这表明,在坡形影响下,只有在生殖生物量较高时生殖配置才会增加,而其他任何参数都无法改变生殖配置大小;坡形条件下生殖配置主要是立地造成的,而不是个体大小造成的。相关分析类似于回归分析,是判断个体与器官大小因素对生殖配置影响一种方法。尽管也用空间代时间取样获得不同大小的个体,但小个体其实是提前衰老个体,而不是幼年旺盛小个体。Li等(2005)研究了固定与半固定沙丘条件下Artemisia halodendron的生殖配置模式,也认识到生殖输出随营养生长而大量增加的现象。但,大小依赖性并不能解释坡形条件下4a年生麻疯树生殖配置的异质性问题,单纯立地能解释生殖配置差异,并且都形成3级等级柱状图(如图2所示)。生殖生物量相对配置与生殖生产力配置在山坳最高,缓坡地、斜坡地、凸坡与凸坡相当,而平地最低,这是水分配置差异造成的。山坳为疏水性凹坡,水分适宜,生殖配置最高。缓坡地林分密度高,斜坡地石块多,凸坡水分限制明显,这些林地生殖配置都相对较低。而平地积水,氧气不足,根系吸收能力不足而根系发育较大而吸收面积增加,这些都限制了生殖器官发育。坡形异质性主导下的资源供给差异可能更能解释麻疯树生殖配置的异质性。
生活史权衡理论能解释这种水分失调状况下坡形主导的生殖配置差异。但在截然异质性环境下,资源限制时水分吸收器官根系资源配置增加,而生殖器官与根系都是接收叶片同化器官资源的汇,生殖配置必然减少,汇间权衡造成了水分限制条件下生殖配置的降低。同时,氧气供应限制也会权衡个体资源配置的方向,使得增加接收面而降低生殖器官的配置。个体发育的资源总是有限的,在营养器官吸收利用器官配置增加必然会降低生殖器官配置比例(张大勇,2004)。生殖配置提高是高产的重要标志,从生活史权衡理论上分析,实际上营养器官过多资源必然会降低生殖输出或生殖配置效率,提高生殖配置只能降低营养器官比重。但营养器官是生殖器官发育的载体,高产必然需要降低间接性维持性大龄营养器官比重,而增加生殖性壮枝培育,以增加生殖器官比重。结合前面麻疯树生殖生物量、种子产量与生物量呈现正相关的结论,减少营养大龄器官配置而增加高生殖力壮枝培育,进而降低营养生长比重而提高生殖器官比重,增加生殖输出与种子生产效率。

表7 不同个体生物量及其配置之间相关分析Table 7 The relation analysis of different individual biomass and organ allocations
群体水平上,麻疯树营养生物量与生殖生物量都受到坡形异质性的塑造,形成了群体生物量的坡形异质性格局。群体生物量与林分密度紧密相关,密度分别为1 111 株·hm-2、1 600株·hm-2、2 500株·hm-2、3 333株·hm-2、5 000株·hm-2、10 000株·hm-2时,总生物量分别达到 18.333 t·hm-2、22.391 t·hm-2、35.887 t·hm-2、36.360 t·hm-2、29.604 t·hm-2、34.458 t·hm-2,种子产量分别为1.246 t· hm-2、2.002 t· hm-2、1.449 t· hm-2、1.153 t·hm-2、0.711 t·hm-2、0.647 t·hm-2(莫丽芬等,2010)。但在密度相对不明显,坡形导致的立地异质性极为明显时,群体生物量差异归结为坡形因素或环境因素,而不是林分因素。在研究麻疯树生殖与生长的土壤厚度差异时推测,土壤因素对生殖的影响是通过根系到营养再到生殖逐级实现的(何亚平等,2010)。卢建国等(2006)研究毛乌素沙地土壤湿度异质性与油蒿种群生物量间关系指出,坡面环境异质性导致了生物量异质性。马店河不同坡形条件自身水文与物质蓄积特性不同,形成了潜在的资源水平异质性,进而增加了根系生物量配置的权衡强度,使得水分或氧气缺乏时根系配置增加进而降低了地上部分资源配置比例,塑造了个体大小、生物量与种子产量异质性。坡形这种个体生物学效应直接决定了群体生物量与种子产量。
马店河栽植麻疯树时间在2005年~2006年,栽植密度为2 100株·hm-2~3 200株·hm-2,群体生物量在2.63 t·hm-2~62.34 t·hm-2之间,群体生产力在1.28 t·hm-2·a-1~37.15 t·hm-2·a-1之间,果实数量在0.63×103个·0.067 hm-2~69.24×103个·0.067 hm-2之间,种子产量在9.21 kg·hm-2~1 472.5 kg·hm-2。莫丽芬等(2010)西双版纳测定的林分密度在1 111株·hm-2~10 000株·hm-2之间,生物量在 18.333 t·hm-2~36.36 t·hm-2之间,种子产量在 0.647 t·hm-2~1.449 t·hm-2之间,栽植时间为2005年。测定林分中凸坡与平地比重不大,不是马店河基地的主体,在斜坡、缓坡与小山坡这类主要坡形下,群体生物量、种子产量分别为 18.98 t·hm-2~62.34 t·hm-2、0.75024 t·hm-2~1.4725 t·hm-2之间。这与西双版纳同龄同密度(2 500株·hm-2~3 333株·hm-2)林分相比,生物量相对较高(vs 35.887 t·hm-2~36.360 t·hm-2),但种子产量相对较低(vs 1.153 t·hm-2~1.449 t·hm-2),但最高产量相当,4 a生麻疯树幼林亩产都能达到100 kg。红河流域麻疯树3 a生幼林生物量为26.025 t·hm-2,净生产力为 10.827 t·hm-2,果实生物量为 2.25 t·hm-2(折合为种子产量为 1.4625 t·hm-2,0.067 hm2产量接近100 kg),与西双版纳、攀枝花麻疯树群体生物量、生产力、种子产量暂时相当。
世界麻疯树分布于降水量从200 mm~1 500 mm 之间时,生产力为1 t·hm-2~10 t·hm-2,栽植5 a后种子产量在0.4 t·hm-2·a-1~1.2 t·hm-2·a-1之间(Openshaw,2000)。Carels(2009)报道了古巴与印度模式种子产量为2 t·hm-2~5 t·hm-2。并且马店河4 a幼林生殖生物量相对配置0.043~0.087之间,生殖相对配置(1与2)为0.07~0.19之间,可能无法实现过大的生殖生产量。只有一种条件下,就是水热充足时高密度幼林促成结实,从结构、立地上突破生殖局限,可能能获得较大的生殖产量。但立足我国麻疯树基地山地自然环境特点,基地内部坡形异质性造成了资源供给异质性,限制了种子产量的全面提高。针对坡形因素的生物生态学效应,凸坡、平地立地改良是必需的,或者改变树种配置(遗传途径),首先提高立地资源度或者物种对立地的适应度,其次优化林分与个体结构配置,寻求特定立地资源高效利用的结构度,实现高产稳产。显然,从系统角度思考,基地种子产量的高产必然与系统斑块的每一个组成的高产,困难立地是经营管理的重点地带;也是经营技术模式运用强度较大的地带。而麻疯树这一树种而言,水分稍微充足时,生长迅速、衰退容易,经营管理对于树体更新显得极其重要。但在干性地区提高适宜而充足的水分,才能提高资源度,经营管理中资源度提高是首要的,也是经营活动实质性与否的标志。
最后,麻疯树耐旱性好,畏惧积水,在水分充足时,根系氧气不足而生长量与积累量最低。避免积水与资源度提高中的保水保墒同等重要。幼树生长期控制水分,长大以后则水分利用能力强大,水分需求增加,利于提高积水性平地的水分利用强度,则积水压力缓解,而水分可能转变为适宜或充足水分。很多麻疯树研究都探讨了耐旱性问题(焦娟玉等,2010),都是阐明树种对水分的适应性,也是抗逆性的一种表现。事实上,种子生产为目标的麻疯树定向培育必然需要在较为充足的水分条件下从事生产,并且提供充足的环境条件来寻求遗传潜能的充分表达。但也需要认识到雨季积水与旱季缺水的水分供应矛盾,以及水分坡形配置异质性现象,科学调度、合理配置,发挥经营者积极性。水分控制局部试验也反应对干热河谷地区水分矛盾的充分认识与协调,如凹坡向凸坡引水、带状水平沟积水保墒、覆草代抚、以耕代抚、平地疏水排水等,还有有机质增施蓄水保墒,这些研究利于协调水分矛盾,也是运用干热河谷水分平衡规律来指导麻疯树高产稳产。这些调控措施也是基于麻疯树水分适应度研究而指导生产实际的。麻疯树基地经营活动也是摆脱自然资源坡形等立地权衡配置格局,而逐渐保蓄调配而达到系统斑块均一优化,在总体上逐渐达到高产稳产所要求的立地资源度、景观结构优化度,最大限度的表达遗传特质与利用环境资源,从而实现生物油料林定向培育高产稳产的基本目标。
[1] 罗建勋,奉正顺,唐平,等.四川小桐子分布特点及适生环境选择初探[J].西南林学院学报,2007,27(3):6 ~10.
[2] 焦娟玉,陈坷,尹春英.土壤含水量对麻疯树幼苗生长及其生理生化特征的影响[J].生态学报,2010,30(16):4460 ~4466.
[3] 郭永清,郎南军,杨旭,等.云南膏桐人工幼龄林生物量和生产力的研究[J].西北林学院学报,2010,25(2):1 -4.
[4] 莫丽芬,唐建维,刀祥生,等.林分密度对小桐子生物量及其产量的影响[J].中南林业科技大学学报,2010,30(4):61 ~68.
[5] Li F R,Zhang A S,Duan S S,Kang L F.Patterns of reproductive allocation inArtemisia halodendroninhabiting two contrasting habitats[J].Acta Oecologica,2005,28,57 ~64.
[6] 卢建国,王海涛,何兴东,等.毛乌素沙地半固定沙丘油蒿种群对土壤湿度空间异质性的响应[J].应用生态学报,2006,17(8):1469~1474.
[7] 何亚平,蔡小虎,费世民,等.土壤厚度对麻疯树生殖与生长性状的影响[J].四川林业科技,2010,31(3):1 ~11.
[8] Openshaw K,2000.A review ofJatropha curcasL.an oil plant unfulfilled promise[J].Biomass Bioenergy,19:1 ~15.
[9] Carels Nicolas,2009.Jatropha curcas:A review[J].Advanses in Botanical Research,50:39 ~86.
[10] 张大勇.植物生殖生态学与生活史进化[M].北京:高等教育出版社,2004.
