川渝地区乌桕天然群体种子性状表型多样性研究
2011-06-26罗建勋辜云杰王红莉
冯 毅,罗建勋* ,杜 波,辜云杰,王红莉,向 青
(1.四川农业大学林学院,四川 雅安 625014;2.四川省林业科学研究院,四川 成都 610081;3.四川龙泉驿区林业局,四川 龙泉驿 610100)
乌桕(Sapium sebiferum(L.)Roxb),大戟科(Euphorbiaceae)乌桕属(SapiumP.Br.)落叶乔木,种子黑色,圆球形,外被白色蜡质假种皮,固着于中轴上。原产于我国,喜光、喜湿、耐热、对土壤要求不严[1]。并且乌桕是集能源,药用,用材,观赏为一体的多功能树种[2],也是我国亚热带重要的木本油料树种。种子既含油又含脂,桕脂可提取类可可脂[3-4],梓油是可转化为生物柴油[5]。现代药理实验表明,乌桕的根、茎、叶具有多种生理活性[6-7]。乌桕木材坚硬致密,纹理清晰,为高档建筑、家具、雕刻和精密模具的良材,枝桠的燃烧值可达17 125 J/g[8]。乌桕树冠圆整,树形优美,深秋时节,树叶变红、黄、橙、紫等色,是著名的彩叶植物。
随着传统用途代用品的逐渐出现,乌桕籽市场销路存在问题,经济效益降低,导致大面积被砍伐,许多遗传资源已丢失[9],急需进行抢救性种质资源的收集和保存。要对乌桕群体进行科学的保护和合理利用,必须弄清种内群体的遗传多样性。表型多样性是遗传多样性研究的重要内容,利用表型性状研究群体的遗传多样性具有简便、快速和节省费用等优点,至今仍然是重要而且有效的。本文报道乌桕种子表型多样性研究结果,主要目的是了解不同地理区域乌桕的种子表型变异程度,揭示其表型变异规律和变异模式。
1 材料与方法
1.1 群体选择与试验材料采集
在川渝地区乌桕的自然分布区,2009年11月份参照苦楝聚合群体抽样方法[10],共收集四川凉山州的会东、雷波,乐山的井研,泸州的古蔺,绵阳的游仙,广元的朝天,达州的万源,重庆的巫山、酉阳等9个有代表性的群体,基本涵盖了该树种在川渝地区的自然分布,并对采样地区的地理生态因子背景值进行了调查(如表1)。群体内单株选择条件参见罗建勋等的研究[11,12]。每群体选取30个单株采样,株间距离50 m以上,在每个单株树冠中上部南向采集100 g以上成熟果实,分单株风干和保存。
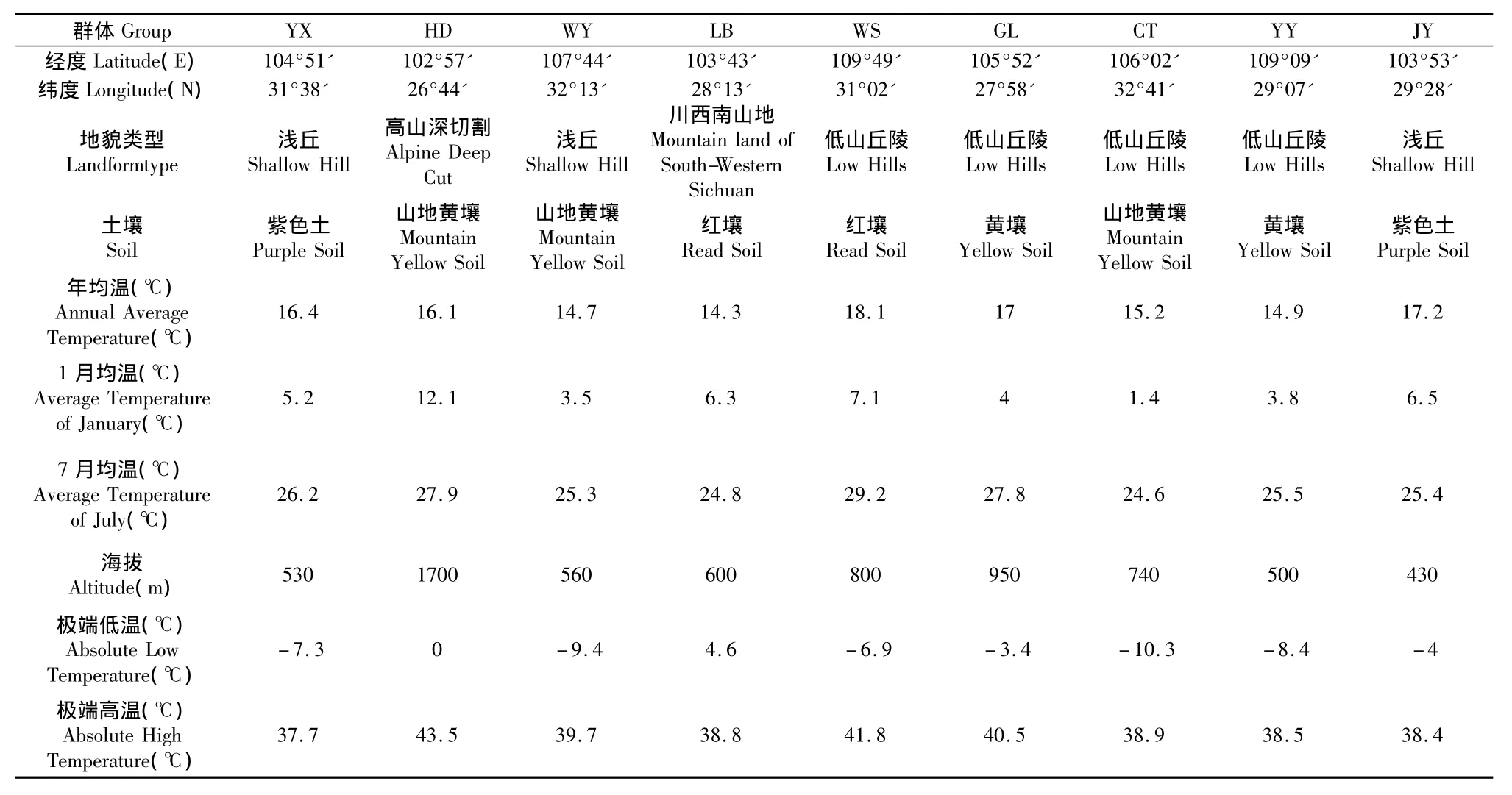
表1 乌桕采集群体的地理位置及气象因子Table 1 The geographical and meteorological factors of Sapium sebiferum groups
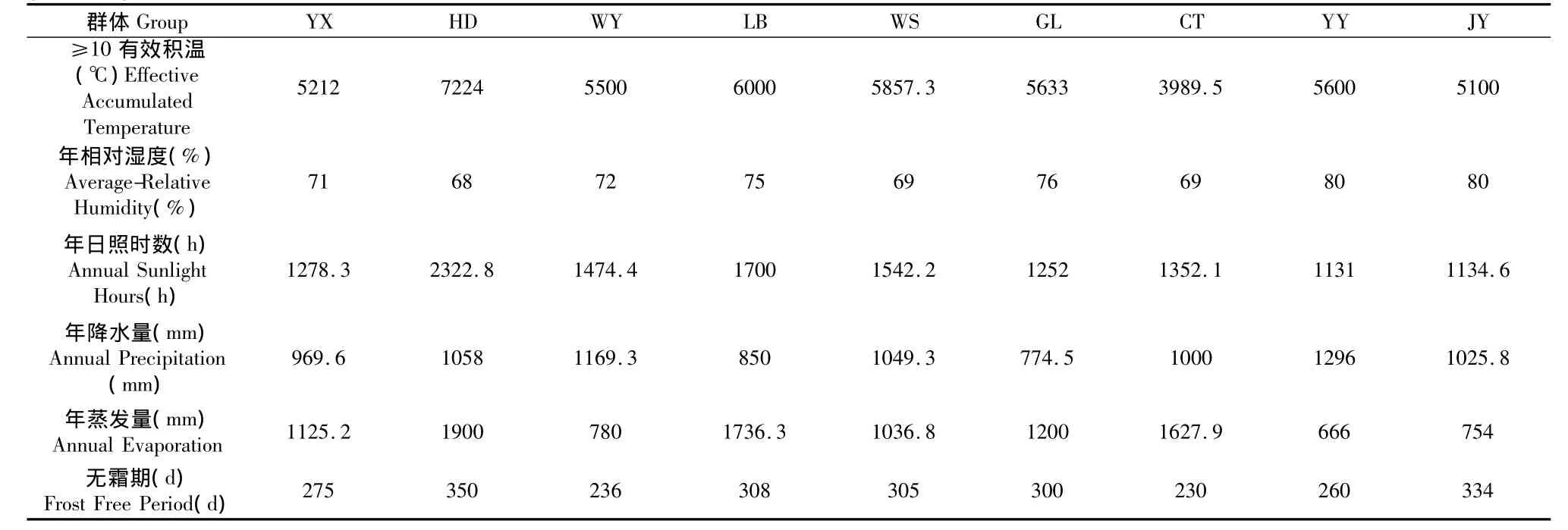
(续表1)
1.2 性状测定方法
未去蜡质层种子,用电子游标卡尺(精度为0.01 mm)测量乌桕种子大小等性状,如长径(平行乌桕种子中轴线最长处)、宽径(垂直种子中轴最宽处)、窄径(种子中轴面与背面的厚度)等;分别计算种子的形状指数,如种子长/宽、种子长/高等;用电子天平称(精度为0.01 g)乌桕种子各重量性状,如单粒重、50粒重等;每个群体测量30个家系,每个家系随机测定50次。测量完成后将所有种子人工去蜡质层后,按照再上述方法再重复测量一次所有种核性状。
1.3 数据处理
巢式设计的线性模型[13-14]为:

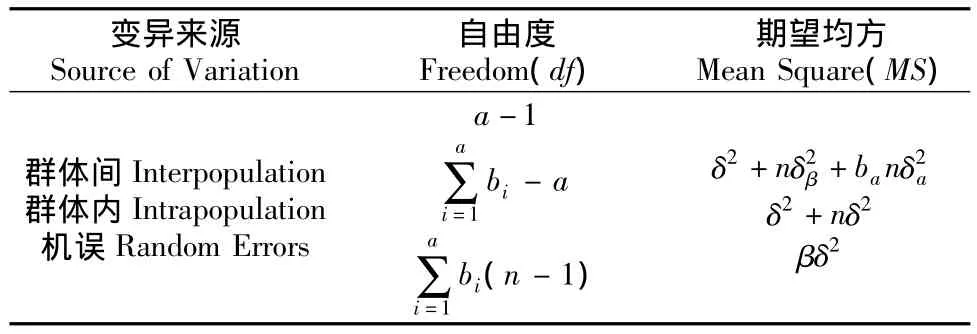
变异来源Source of Variation自由度Freedom(df)期望均方Mean Square(MS)群体间Interpopulation群体内Intrapopulation机误Random Errors a-1∑a i=1 bi-a∑a i=1 bi(n-1)δ2+nδ2 β +banδ2a δ2+nδ2 βδ2
式中:i=1,2,…,a;j=1,2,…,bi;k=1,2,…,n;μ-总平均值;τi-群体间效应值;βj(i)-群体内家系效应值;εk(ij)- 第 ijk 个观测值的误差分别为群体间、群体内、机误的方差分量。
个体重复力[15]计算公式:R=(MSB-MSGW)/[MSB+(k-1)MSGW]
式中:R-重复力,MSB-组间均方,MSGW-组内均方,k-组内个体数
群体间表型分化系数[16-17]计算公式
式中:V-表型分化系数,表示群体间变异占遗传总变异的百分比.
采用Excel和SPSS 10.0[18]统计软件对各性状进行描述性统计、方差分析、Duncan多重比较、相关性分析及聚类分析。
2 结果与分析
2.1 乌桕群体间的形态变异特征
乌桕群体间和群体内表型性状的变异均方及F值见表2。经F检验,种子长、种子宽等种子的16个表型性状在群体间和群体内都存在显著差异。16个表型性状均值及均值的多重比较见表3。从表3可以看出,乌桕种子的形态特征在群体间存在极显著差异,LS、WS、HS和50SGW的变异特征都是北部WY群体最小,SKW为CT群体最小但与WY群体差异不显著,LS以WS群体最大与YY群体无显著差异,WS以LB群体最大与WS群体无显著差异,HS以LB群体最大,SKW与种子50SGW均为WS群体最大,LWS以YX群体最小与HD群体无显著差异,LHS以HD群体最小,LWS和LHS均为CT群体最大。SKW与50SGW均为WY群体最小,WS群体最大;去蜡质层后:LSN、WSN、HSN、SKWQ、50SGWQ均是LB群体最大,LWSN以GL群体最大,LHSN、50WSGW和50WASP均为WS群体最大;LSGW最小是WY群体,WSN、HSN、SKWQ和50SGWQ最小是 CT群体;LWSN最小是 YY群体,LHSN、50WSGW和50WASP最小均是HD群体。
以上结果表明,LS、WS等表示种子大小的性状与重量性状随着纬度的增加逐渐减小,综合起来WY、CT群体性状较差,WS、YY、LB群体性状较优。
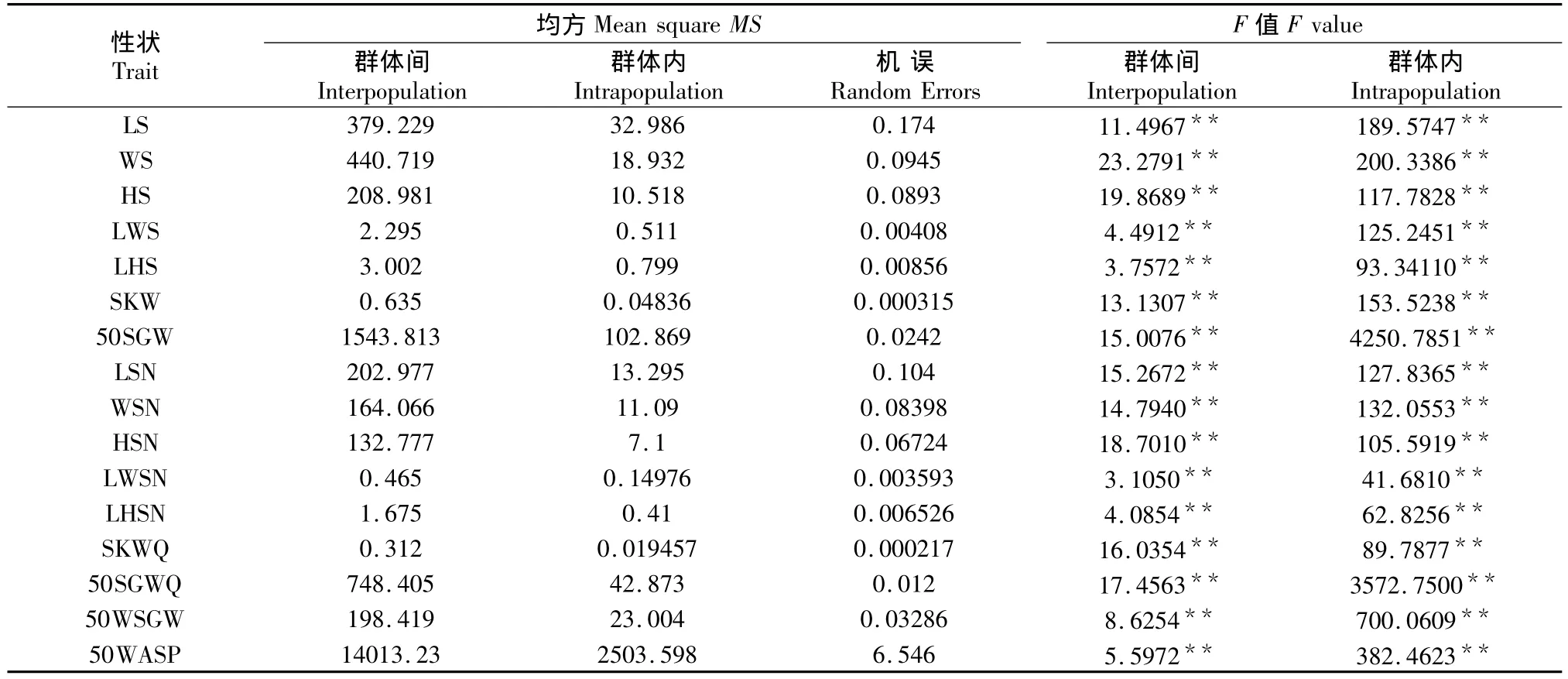
表2 乌桕天然群体间群体内表型性状的方差分析结果Table 2 Variance analysis of interpopulation and intrapopulation phenotypic traits of S.sebiferum

表3 乌桕天然群体间表型性状的平均值及标准差Table 3 The mean and standard deviation of phenotypic traits of S.sebiferum Interpopulations
2.2 乌桕群体内的形态变异特征
用变异系数表示性状值离散性特征,变异系数越大,则形状值离散程度越大。乌桕9个群体表型性状的变异系数见表4。从表4可以看出乌桕种子16个性状总变异系数变幅为6.84% ~39.54%;但是,乌桕种子形状大小等性状变异系数的变幅为6.84%~11.86%,说明种子性状大小等性状稳定性高;乌桕种子的各重量性状的变异系数均超过20%;50WSGW的变异系数更是高达39.54%,说明乌桕种子的各重量性状的稳定性相对较低,选择潜力较大,而这些性状正好是乌桕经济价值的重要形状。从表4还可以看出不同群体内的每一性状存在一定的差异,说明不同群体的环境异质性导致了群体表型变异的差异。WY和CT群体平均变异系数较小,这可能和WY和CT群体正好是乌桕天然分布的北界有关;YX群体变异系数最大(18.42%),这可能预示YX群体是乌桕在川渝地区的表型多样性分布中心。
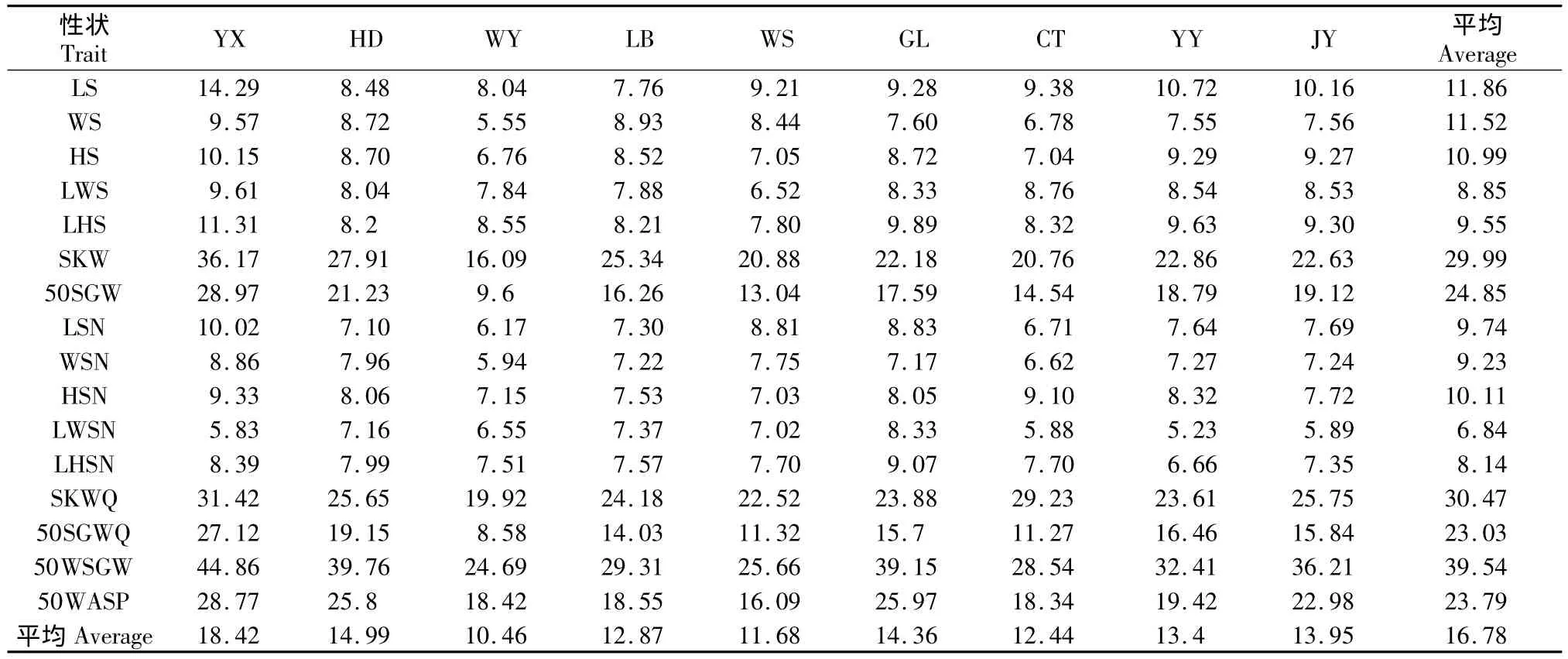
表4 乌桕9个天然群体表型性状的变异系数(%)Table 4 Coefficients of variation of phenotypic traits of the 9 S.sebiferum groups(%)
2.3 桕种子性状的重复力
重复力是基因型和一般环境的共同作用结果,可以消除特殊环境作用和误差的影响[16],由于一般环境的影响在同一个群体中变化不大,因此重复力也可以作为广义遗传力的估计上限。所以重复力的大小在一定程度上能代表某个群体某一性状的遗传稳定性。从表5可以看出,9个群体的50SGW平均重复力为(0.9775),YX群体最高达到(0.9913),WS最低也达到(0.9580)。50SGWQ和50WSGW以及50WASP的重复力均超过0.8,说明作为乌桕最重要的几个经济性状均具有很高的稳定性,具有很高的选择价值。群体平均重复力从大到小依次为,YX>GL>JY>YY>CT>HD>LB>WY>WS。各种子性状的平均重复力从大到小依次为50SGW>50SGWQ>50WSGW>50WASP>LS>WS>SKW>LWS>LSN>WSN>HS>SKWQ>LWS>HSN>LHSN>LHSN。
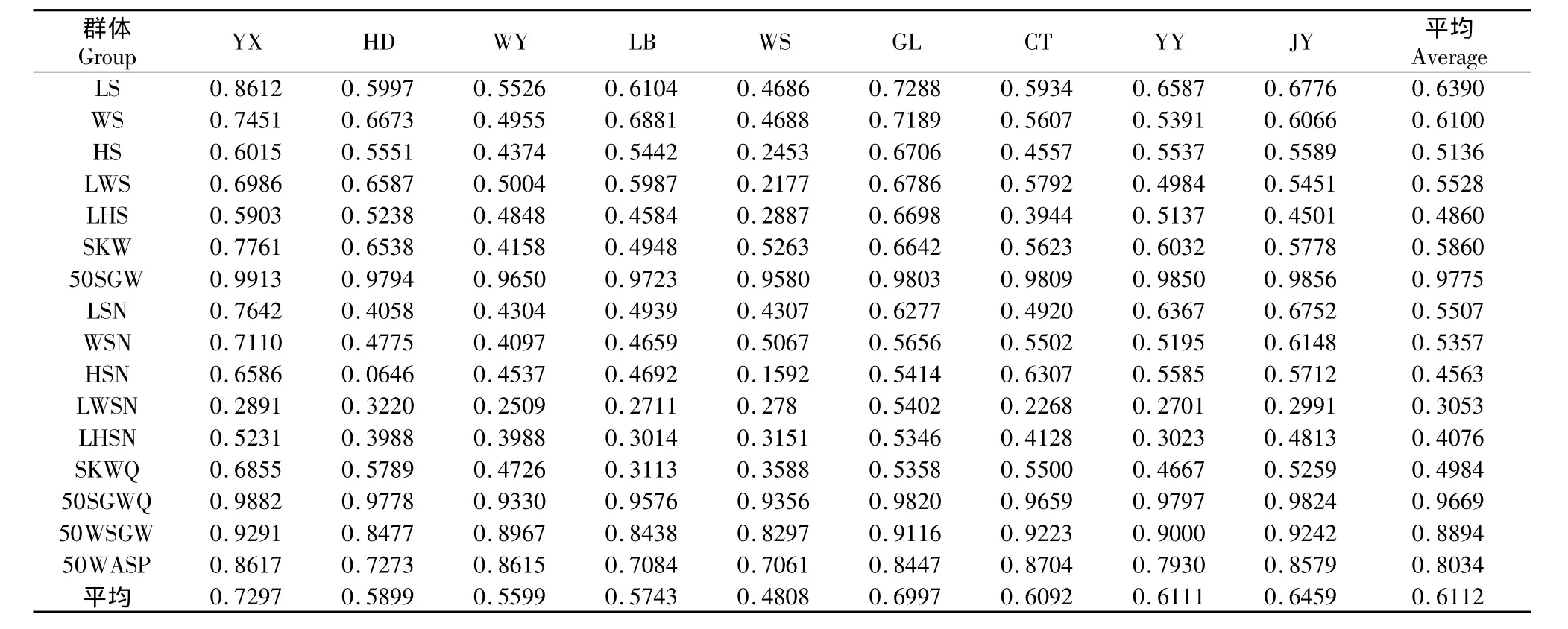
表5 乌桕9个群体表型性状的重复力Table 5 Repeatability of the phenotypic traits of the 9 S.sebiferum groups
2.4 乌桕天然群体间表型分化
按巢式设计方差分量比组成了各方差分量占总变异的比例(表6)。从表6可以看出种子各性状的表型分化系数变异幅度为6.7069%~42.7381%,其中去蜡质层和未去蜡质层的各大小形状和重量性状的表型分化系数均超过平均值,说明乌桕种子各大小形状和重量性状在群体间的变异较大,但去蜡质层和未去蜡质层的各种子形状指数的表型分化系数明显低于平均值,说明种子形状指数在群体间的变异较小,总体来说种子形状较为稳定,平均表型分化系数为25.4732%,说明乌桕种子群体内多样性(74.5268%)显著高于群体间的多样性(25.4732%)。

表6 乌桕表型性状的方差分量及表型分化系数Table 6 Variance portions and differentiation coefficients of phenotypic traits of intrapopulation and interpopulation of S.sebiferum
2.5 乌桕种子表型性状相关分析
表7对乌桕种子16个表型性状进行相关分析与检验,结果表明乌桕 WS、HS、LSN、WSN、SKWQ、50SGWQ均与其余11个性状存在显著相关,LS、HSN、SKW、50SGW均与其余10个性状存在极显著相关,50LE与其余9个性状显著相关,LWS与其余8性状显著相关。表明 LS、WS以及 SKW 和50WSGW是乌桕最重要的表型性状。

表7 乌桕种子表型性状间相关关系矩阵Table 7 The correlation analysis matrix between each phenotypic trait
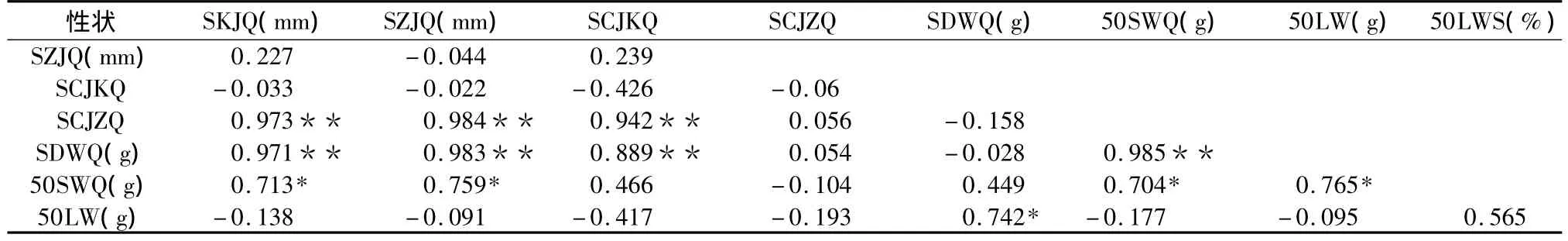
(续表7)
2.6 乌桕种子性状和采集点的地理生态因子的相关关系
将乌桕杉的16个种子性状与采样点的地理生态因子进行相关分析与检验(表8):LWS、LHS分别与纬度纬度呈显著正相关,分别与1月均温、极端低温、有效积温和无霜期呈显著负相关;HS与纬度和无霜期呈显著负相关;HSN与纬度呈极显著负相关,与极端低温和无霜期呈显著正相关;LWSN与降雨量呈极显著负相关;LHSN与纬度显著正相关,与1月均温、海拔、有效积温、日照时数分别呈显著负相关;SKWQ与无霜期呈显著正相关。结果表明:乌桕种子性状的变异主要是以纬度和温度变异为主,即随着纬度的增加、极端低温的降低、1月均温的降低、有效积温的减少、无霜期天数的减少乌桕的种子都会从长圆核型向长核型逐渐变异;水分和光照对种子性状也有一定的影响,随着日照和降雨量的增加,乌桕去蜡质层后的种子也呈现愈来愈饱满的趋势;但均无温度影响明显,在温度的影响中又以低温影响为主,高温对其影响不大,这说明低温可能是影响乌桕种子性状的主要因子。

表8 乌桕种子性状和采集点的地理生态因子的相关关系Table 8 Correlation coefficients between the seed traits and geo-ecological factors based on the populations of S.sebiferum
2.7 乌桕群体表型聚类分析
利用欧式平均距离,采用类间平均连锁法对9个群体的16个种子表型数据进行聚类分析,结果见图1。以欧氏距离7为阈值,9个群体明显分为4组。北部的WY群体与CT群体首先聚在一起再与WS群体聚为一组,LB群体和YY群体聚为一组,YX群体首先JY群体聚在一起再与GL群体聚为一组,最南端的HD群体单独聚为一组。

图1 乌桕种子性状聚类图Fig.1 UPGMA cluster of the seed traits of S.sebiferum
3 结论与讨论
3.1 川渝地区乌桕种子变异规律和变异模式
对形态性状变异的度量、描述和分析是探讨进化问题的基础[19]。形态性状既具有变异性又具有稳定性,受其本身的遗传组成和所处生态环境两方面的影响,是生物适应其生存环境的表现形式[20]。乌桕各形状的变异随着纬度的增加、极端低温的降低、1月均温的降低、有效积温的减少、无霜期天数的减少乌桕的种子都会从长圆核型向长核型逐渐变异。其次,随着日照和降雨量的增加,乌桕去蜡质层后的种子也呈现愈来愈饱满的趋势。高温对其无影响,进一步印证了乌桕喜光,喜湿,喜热不耐寒,说明乌桕种子变异主要是受低温影响,其次是光照和湿度。
3.2 乌桕遗传改良策略讨论
乌桕种子表型形态特征的多样性往往具有适应意义,天然群体保持较大的多样性对植物本身而言是有利的。乌桕在我国长江以南广泛分布,从近海平面到海拔2 800 m均有分布,垂直跨度大,且水系不同,环境条件复杂,微立地效应十分明显,通过长期的地理隔离,自然选择和人工选择,使得乌桕产生了极其丰富的种类变异。川渝地区的乌桕种子性状在群体间、群体内差异均达到极显著。不同性状的变异系数变幅为 6.84% ~39.54%,平均为16.78%;不同性状的重复力变幅为0.3053~0.9775,平均为0.6112。种子形状指数各性状变异系数均小于种子重量各性状变异系数,重复力亦是如此,说明在对乌桕进行遗传改良时,种子形状指数各性状不仅变异系数小,而且重复力低(重复力是广义遗传力的估计上限,在一定程度上可代替遗传力),改良选择难度较大;种子各重量性状刚好相反,所以应加大对种子各重量性状的选择;表型变异必然蕴藏着遗传变异,表型变异越大,可能存在的遗传变异越大。根据乌桕丰富的自然变异情况,可以知道乌桕的改良前景是广阔的。川渝地区乌桕群体内分化高于群体间,该研究与大戟科另一能源树种麻疯树研究结果[21~23]相似,因此,在遗传改良工作中,在进行优良种源、优良家系选择和利用的同时,应加大优良个体选择和利用的力度。
3.3 乌桕现有天然资源之中种质资源保存策略探讨
本文所研究的均是表型变异,是遗传型和环境因子共同作用的结果,因此其更具有综合性和实用性。通过对乌桕种子的遗传多样性及群体遗传结构的研究,提出其以原地原地保存为重点,异地保存相结合的保存策略:原地保存策略[24]:首先原地保存应具有典型性,由变异系数得知,遗传多样性的丰富度依次为 YX、HD、GL、JY、YY、LB、CT、WS、WY,再综合乌桕群体间聚类的遗传距离,因此确定乌桕天然群体原地保存抽样群体为YX群体、HD群体、YY群体、WS群体;异地保存应具有广泛性,在异地保存中,要尽可能保存基因的多样度,因此应尽可能多地收集其他地区种质材料,并且不断补充,不断丰富种质资源库。在重视不同群体的保存时,同时兼顾群体内不同个体的保存,为进一步开发利用乌桕的遗传变异打下基础。
[1] 金代钧,黄惠坤.乌柏的地理分布和环境的关系[J].广西植物,1984,4(1):71 ~80.JIN D J,Huang H K.The Distribution ofSapium sebiferumRoxb.In Relation to the Environmental Conditions[J].Guihaia,4(1),71~80.
[2] 李冬林,黄 栋,王 瑾,金雅琴等.乌桕研究综述[J]江苏林业科技2009,36(4):43~47.LI DL,HUANG D,WANG J.The review of research onSapium sebiferum[J].Journal of Jiangsu Forestry Science and Technology,2009,36(4):43 ~47.
[3] 喻艳,程晶,宋峥,等.从乌桕脂中提取类可可脂方法的改进[J].化学与生物工程,2004,35(3):48 ~49.YU Y,CHENG J,SONG Z.Improvements of Method on Extracting Cocoa Butter Equivalent fromSapium sebiferumButter[J].Chemistry and Bioengineering,2004,35(3):48 ~49.
[4] 李晓航,乌桕类可可脂在巧克力生产中的应用[J].林产化工通讯,1992,(2):26 ~28.LI X H.WuJiu class cocoa butter in chocolate production application[J].Journal of Chemical Industry of Forest Products,1992(2):26~28.
[5] 杨志斌,齐玉堂,王晓光,等.乌桕籽制取生物柴油研究初报[J].湖北林业科,2007(6):32 ~34.YANG ZB,QI YT,Wang XG.Preliminary Report on Research ofSapium sebiferumRoxb.Seed Producing Biodiesel[J].Hubei Forestry Science and Technology,2007(6):32 ~34.
[6] 郭晓庄.有毒中药大辞典[M].天津:天津科技翻译出版公司,1991.GUO XZ.Toxic medicine dictionary[M].Tianjin:Tianjin science and technology translation and publishing company,1991.
[7] Hsu F L,Lee Y Y,Cheng J T.Antihypertensive activity of 6-O-galloyl-D-glucose,a phenolic glycoside from Sapium sebiferum [J].Nat Prid,1994,57(2):308 ~312.
[8] 王育民,汪企明,王新,等.江苏省61个树种木材燃烧热的测定[J].江苏林业科技,1992,19(2):6 ~12.WANG YM,WANG QM,WANG X.Determination of combustion heat for 61 tree species in Jiangsu[J].Journal of Jiangsu Forestry Science & Technology,1992,19(2):6 ~12.
[9] 张克迪 林一天.中国乌桕[M]中国林业出版社,1991.ZHANG KD,Lin YT.Sapium sebiferumof China[M].China's forestry press,1991.
[10] 程诗明.苦楝聚合群体遗传多样性研究与核心种质构建[D].中国林科院,2005.CHEN SM.Study on Genetic Diversity of Multitudinous Populations and Construction of Core Germplasm in Melia Azedarach[D].Chinese Academy of Forestry,2005.
[11] 罗建勋,顾万春.云杉天然群体表型多样性研究[J].林业科学,2005,41(2):66 ~72.LUO JX,GU WC.Study on Phenotypic Diversity of Natural Population inPicea asperata[J].Scientia Silvae Sinicae,2005,41(2):66~72.
[12] 辜云杰,罗建勋,吴远伟.川西云杉天然种群表型多样性[J].植物生态学报,2009,33(2),291 ~301.GU YJ,LUO JX,WU YW.Phenotypic Henotypic Diversity in Natural Populations ofPicea balfourianain Sichuan,China[J].Journal of Plant Ecology(Chinese Version),2009,33(2),291 ~301.
[13] 李斌,顾万春,卢宝明.白皮松天然群体种实性状表型多样性研究[J].生物多样性,2002,10(2):181 ~188.LI B,GU WC,LU BM.A study on phenotypic diversity of seeds and cones characteristics inPinus bungeana[J].Chinese Biodiversity,2002,10(2),81 ~188.
[14] 李梅,韩海荣,康峰峰,等.辽东栎天然群体表型多样性研究[J].北京林业大学学报,2005,27(5):10 ~16.LI M,HAN HR,KANG FF.orphologic variation of leaves ofQuer-cus liaotungensisKoidz.in Lingkong Mountain,Shanxi Province[J].Journal of Beijing Forestry University,2005,27(5):10 ~16.
[15] 续九如.林木数量遗传学[M].高等教育出版社,2006.XU JR.Trees quantitative genetics[M].Beijing:Higher Education Press,2006.
[16] Halil M A K.Genetic of cone morphology of black spruce(Picea mariana(Mill).B.S.P)in Newfoundland Canada[J].Silvae Genetiea1984,33(1):101 ~109.
[17] Halil M A K.Genetic variation in eastern white spruce(Picea glauca(Moeneh)Voss)populations.Can J For Res1985,15(2):444~452.
[18] 三味工作室.SPSS v10.0 for windows实用基础教程[M].北京:北京希望电子出社,2002.三味工作室.SPSS v10.0 for windows Practical foundation course[M].Beijing:Beijing hopes to electronic publishing house,2002.
[19] 葛颂,洪德元.泡沙参复合体(桔梗科)的物种生物学研究:Ⅲ.性状的遗传变异及其分类价值[J].植物分类学报,1995,33(5):433~443.GUO S,HONG DY.Biosystematic studies onAdenophora potaniniiKorsh.Complex(Campanulaceae)Ⅲ genetic variation and taxonomic value of morphological characters.Acta Phytotaxonomica Sinica,1995,33(5):433 ~443.
[20] 杨继.植物种内形态变异的机制及其研究方法[J].武汉植物学研究1991,9(2):185~195.YANG J.Infeaspecific variation in plant and the exploring methods[J].Journal of Wuhan Botanical Research,1991,9(2):185~195.
[21] 辜云杰,何朝均,张俊等凉山州麻疯树天然群体种子性状多样性研究[J].西南林学院学报2009,29(6),11 ~14.GU YJ,HE CJ,ZHANG J.Study on Phenotypic Diversity of Jatropha curcas Seeds from Natural Populations in Liangshan Yi Autonomous Prefecture.Journal of Southwest Forestry University,2009,29(6):11 ~14.
[22] 罗建勋,和献峰,辜云杰,等.攀枝花地区麻疯树天然群体表型多样性研究[J].西南林学院学报,2008,28(6):31 ~35.LUO JX,HE XF,GU YJ.Study on Phenotypic Diversity of Natural Populations ofJatropha curcasin Panzhihua Area,Sichuan Province.Journal of Southwest Forestry University,2008,28(6):31~35.
[23] 向振勇,宋松泉,王桂娟,等.云南南部不同种源地小桐子遗传多样性的ISSR分析[J].云南植物研究,2007,29(6):619~624.XIANG ZY,SONG SQ,WANG GJ.Genetic Diversity ofJatropha curcas(Euphorbiaceae)Collected from Southern Yunnan,Detected by Inter- simple Sequence Repeat(ISSR).Acta Botanica Yunnanica,2007,29(6),619 ~624.
[24] 宋丛文,包满珠.[J].植物生态学报,2005,29(3):422 ~428.SONG CW,BAO MZ.Sampling Strategy For Preservation of Germ Plasm fromDavidia involucrata[J].Journal of Plant Ecology(Chinese Version),2005,29(3):422 ~428.
