烤烟易烤性遗传分析
2011-06-14王英俊翟明泉徐秀红张兴伟张忠锋
王英俊,倪 超,翟明泉,徐秀红,张兴伟,张忠锋,*
(1淄博烟草有限公司 ,山东淄博 255035;2川渝中烟工业公司,四川成都 610016;3中国农业科学院烟草研究所,农业部烟草类作物质量控制重点开放实验室,山东青岛 266101)
近年来,在大力发展中式卷烟的同时,国家烟草专卖局又进一步提出“卷烟上水平”的行业目标。烟叶作为行业的基础性物质,被放在了十分重要的位置。而田间收获的鲜烟叶,必须经过烘烤加工才能调制成符合卷烟工业需求的原料。烘烤是烤烟生产中的一个重要环节,烤烟品种的烘烤特性与遗传基础、栽培条件、烘烤工艺等有关,其中,品种被公认为是影响烟叶烘烤特性的最重要因素[1]。烘烤特性包括易烤性和耐烤性两方面。易烤性反映烟叶在烘烤过程中变黄、脱水的难易程度及同步程度,主要反映烟叶的变黄特性[2]。可以用烟叶的变黄特性(变黄指数[3])来评价烤烟个体的易烤性。
植物数量性状主基因+多基因混合遗传模型分析法[4]可检测和鉴定数量性状主基因和多基因的存在,并估测其遗传参数。目前该方法已在多种作物的多个数量性状的遗传研究中得到成功应用[5~10]。在烟草的株高性状[11]上也已经成功应用,但在烤烟品种易烤性遗传研究上的应用尚未见报道。
本研究以中烟100(易烤性好的烤烟品种)和翠碧1号(易烤性相对较差的烤烟品种)[12]为亲本构建了 P1,P2,F1,B1,B2和 F2等6个世代群体,以苗期烟叶的变黄指数作为易烤性指标,利用数量性状主基因+多基因混合遗传模型,对烤烟品种易烤性的遗传效应进行了初步研究,旨在揭示烤烟品种亲子代间易烤性性状的遗传关系,为烟草烘烤和育种的定向改良提供理论依据。
1 材料与方法
1.1 材料
将易烤性相对较好的烤烟品种中烟 100和易烤性相对较差的烤烟品种翠碧 1号分别作为亲本 P1,P2,杂交获得 F1,F1自交获得 F2,F1×P1获得 B1,F1× P2获得B2,从而组成6个世代群体。上述材料均由中国农业科学院烟草研究所提供。
1.2 方法
在中国农业科学院烟草研究所温室中播种中烟100、翠碧 1号及其 F1,B1,B2和 F2的 6个世代群体。小十字期进行假植,4~5片真叶时移栽到直径 13~14 cm的小花盆中,第 8片真叶长出后打顶,留叶8片。两周后,自上而下采收 3片叶,以报纸包成面包(书本)状,放置在温度 36℃、相对湿度 90%的恒温恒湿箱内,每24 h测定一次变黄比例,累计测7次,算出变黄指数(Y I)。
YI=∑ Y/n,其中 n为测定次数,Y为各次的变黄比例(3片叶的平均值)。YI值愈大,变黄愈快,表示易烤性愈好。
各供试材料的管理措施同常规方法,其生长环境条件基本一致。亲本、F1各测量了20株,B1,B2和F2分离群体各测量了 193,202和 196株。
1.3 数据分析
采用植物数量性状主基因+多基因混合遗传模型多世代联合分析方法,对上述 6个世代群体的变黄指数进行联合分析,通过极大似然法和 IECM(iterated expectation and conditional maximization)算法对混合分布中的有关成分分布参数做出估计,然后利用AIC(Akaike′s information criterion)准则和一组适合性测验,选择最优遗传模型,并估计主基因和多基因效应值、方差等遗传参数[13]。适合性检验共有 5个统计量,即 U21,U22,U23,nW2和 Dn,其中 U21,U22,U23为均匀性检验,nW2为Smirnov检验,Dn为Kolmogorov检验。主基因遗传率h2mg、多基因遗传率h2pg、势能比值 Rp和显性程度Dd的计算按参考文献[14]进行。
2 结果与分析
2.1 变黄指数测定值的次数分布
各世代变黄指数的次数分布列于表1。P1,P2的平均值分别为 24.70和 9.60,F1的平均值为 13.09,介于两亲本之间,并偏向于不易烤亲本 P2。B1,B2和 F2的平均值为 15.38,10.66和 12.80,且变黄指数多集中在7%~ 14%。从各分离世代的数值分布区间,可以看出B1,B2和 F2世代都呈单峰较明显的偏态分布,变黄指数的遗传表现出主基因的特征,进而说明烤烟品种易烤性的遗传可能受主基因控制。
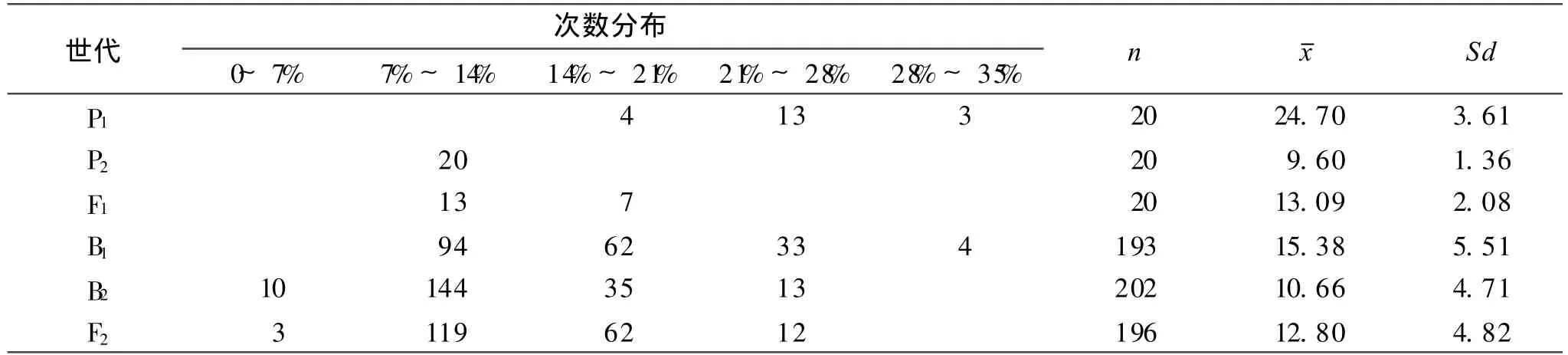
表1 变黄指数次数分布Table 1 Frequency distribution of yellowing index
2.2 遗传模型的分析
用主基因+多基因混合遗传模型的多世代联合分析法对本试验 6个世代群体的易烤性指标变黄指数进行分析,通过 IECM算法,获得 1对主基因 (A)、2对主基因(B)、多基因(C)、1对主基因+ 多基因 (D)和 2对主基因+多基因(E)共5类24种遗传模型的极大似然函数值和 AIC值(表2)。根据 AIC准则,在备选遗传模型中,AIC值最小者可能为最优模型。
由表2可得出,AIC值较低的是 E0,E1,E3和 D1共 4个模型,其 AIC值分别是 3739.81,3746.16,3750.06,3755.61。初步选取这4个模型作为本试验品种的适合性模型。其中,E0为 2对加性-显性-上位性主基因+加性-显性-上位性多基因模型;E1为 2对加性-显性-上位性主基因+加性-显性多基因模型;E3为 2对加性主基因+加性-显性多基因模型;D1为 1对加性-显性主基因+加性-显性多基因模型。
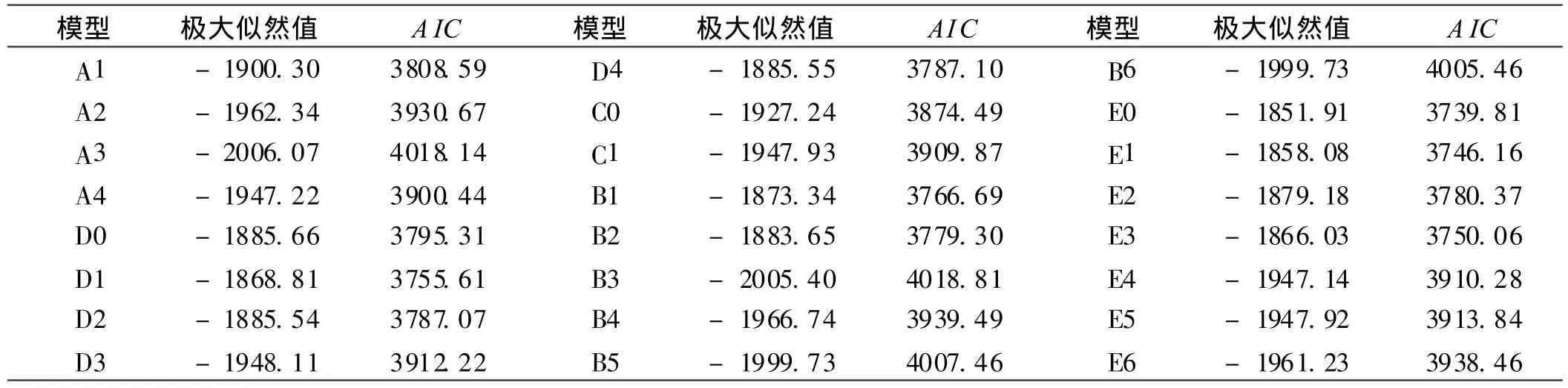
表2 各模型的极大似然值及 AIC值Table2 Maximum likelihood value and AIC value of various models
对这4个备选中两个最适合模型 E0和E1,进行适合性检验(表3),选择统计量达到显著水平个数最少的模型作为最优模型。统计结果表明:模型 E0中有3个统计量与该模型的差异达到显著水平;E1模型中有 2个统计量与该模型的差异达到显著水平,故确定 E1模型为最优遗传模型。初步分析确定,烤烟品种易烤性的遗传可能是由 2对加性-显性-上位性主基因+加性-显性多基因混合控制。
E1模型的全部一阶、二阶分布参数估计值均列于表 4,由表 4可以计算出一阶、二阶遗传参数估计值[3]。
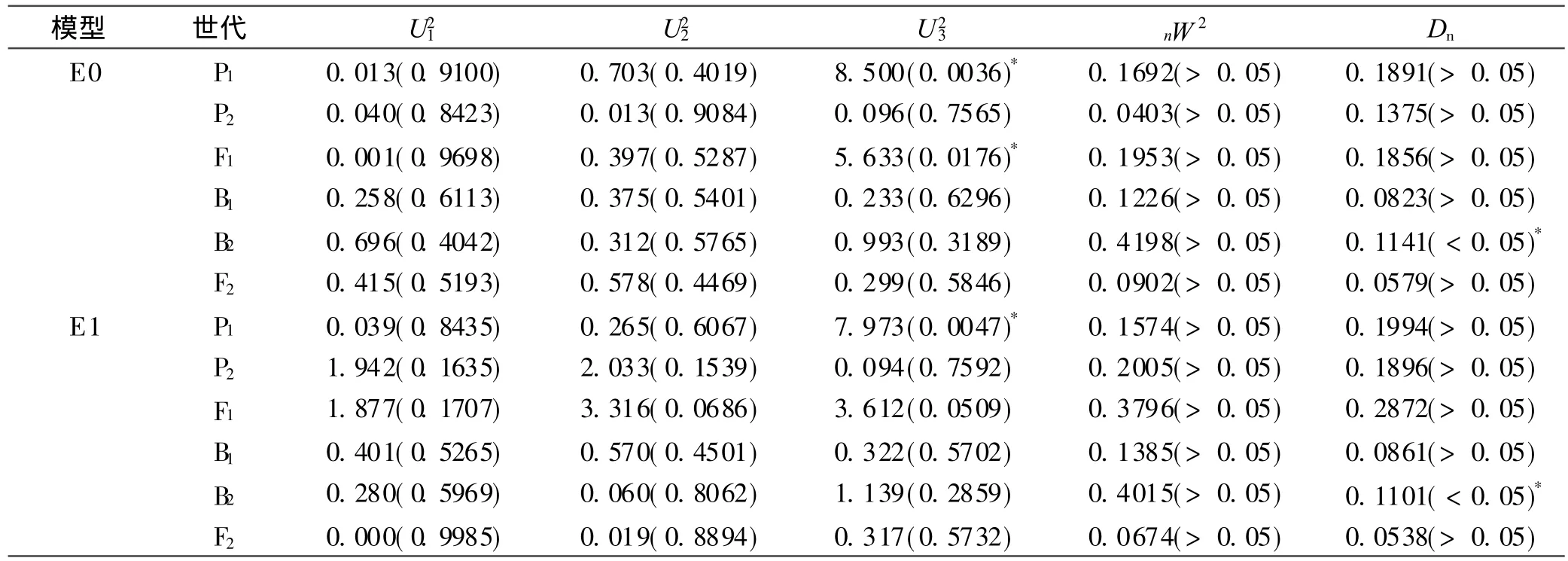
表3 备选模型的适合性检验Table 3 Fitness test of alternative models
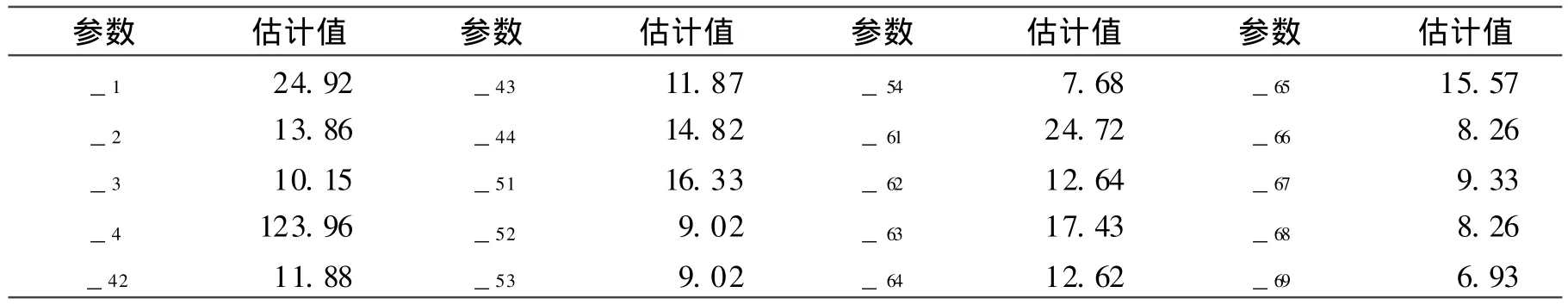
表4 E 1模型成分分布参数的极大似然估计值Table 4 Maximum likelihood value of composition distribution of E1model
2.3 遗传参数估计
E1模型的各成分遗传参数估计值列于表5。本组合6世代变黄指数2对主基因的加性效应均为正值,其中大基因加性效应为6.47,次大基因加性效应为2.42,说明大基因对于加性遗传的贡献率较大;而两者显性效应近似相等,分别为-4.16和-4.15,均为负向效应,说明两对主基因均表现为不完全显性。大基因的加性效应值大于显性效应值,而次大基因的显性效应值大于加性效应值,说明大基因存在正向加性效应,且存在不太强的负显性效应;次大基因存在较强的负向显性效应。
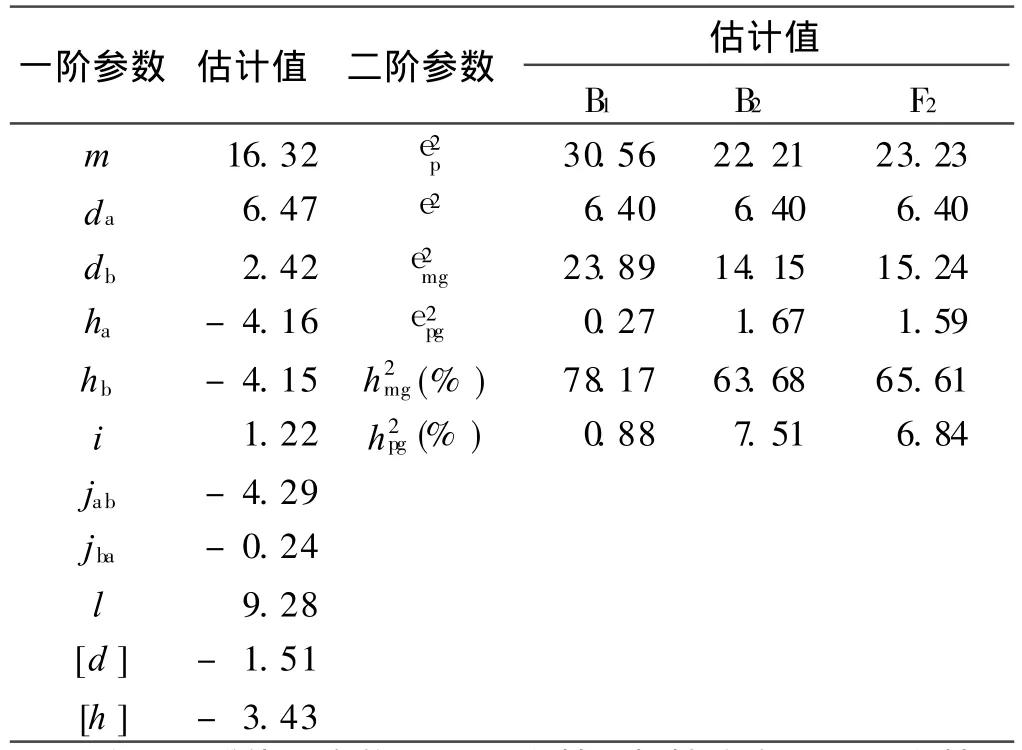
表5 E 1模型变黄指数的遗传参数估计值Table5 Estimates of genetic parameters of yellowing index in E1model
两对主基因之间的加性×显性效应为-4.29,显性×加性效应为-0.24,说明 2对主基因间存在一定的负向互作效应,且互作效应以加性为主。两对主基因间显性×显性效应为 9.28,说明两对主基因间存在较强的杂种优势遗传效应。而两基因间的加性×加性效应不甚明显,为1.22。多基因加性效应与显性效应均表现为负向性,分别为-1.51和-3.43,负向显性效应为主。
通过同质群体所提供的环境误差方差的无偏估计,并对二阶遗传参数进行估计得出,本组合主基因遗传率 B1最高,达 78.17%,B2与 F2相差不大,分别为63.68%和 65.61%,说明本试验烤烟品种的易烤性指标变黄指数的遗传特点表现出较高的主基因遗传效应。多基因遗传率 B2最高,为 7.51%;其次是 F2,为6.84%,B1最低,为0.88%。主基因+多基因效应决定了各分离世代易烤性性状变异的 71.19%~79.06%。
3 讨 论
烤烟作为一种吸食性叶用经济作物,烘烤特性的好坏是衡量其烟叶质量的重要指标。田间收获的鲜烟叶必须经过烘烤才能固定和体现其优良品质,成为商品烟叶[15]。影响烟叶烘烤特性的因素很多,其中,遗传基础(品种)被公认为影响烟叶烘烤特性的最重要因素。日本的藤田茂隆等通过经典遗传学方法研究表明,烤烟品种的易烤性,可能受一个主效基因所支配[3]。本研究利用数量遗传模型的研究结果与之不同。本研究利用P1,P2,F1,B1,B2和F2这6个世代群体对易烤性表型进行联合分析,增加了试验的精确度,提高了主基因遗传参数的估计精度,保证了分析结果的准确性,而且还可以鉴别多基因的存在。
应用于改变任何特定品质性状的育种战略是由控制该性状的基因数目确定的[16]。对于烟草烘烤特性这样复杂的性状,估计涉及的基因数目是很多的,不能简单地用孟德尔方式分析推断性状的遗传组成。烘烤特性是烟草十分重要的性状,但是目前,国内乃至国际对于烟草烘烤特性遗传规律的研究甚少,故有必要对其开展研究。因此,需要在已有研究基础上,对烤烟品种的易烤性以及整体烘烤特性的遗传规律继续深入研究。随着遗传学研究技术和手段的不断提高,烤烟品种易烤性的遗传效应将会进一步明确。
4 结 论
本研究利用植物数量性状主基因+多基因混合遗传模型分析结果表明,烤烟品种的易烤性性状的遗传,符合2对加性-显性-上位性主基因+加性-显性多基因混合遗传模型(E1)。 2对主基因间存在一定的负向互作效应,且互作效应以加性为主。主基因遗传率以B1最高,达78.17%,B2与 F2相差不大,分别为63.68%和 65.61%,表现出较高的主基因遗传效应;多基因遗传率 B2最高(7.51%),其次是 F2(6.84%),B1最低(0.88%)。主基因+多基因效应决定了各分离世代易烤性性状变异的 71.19%~79.06%。
[1]唐经祥,孙敬权,任四海.烤烟不同品种烘烤特性的研究初报 [J].安徽农业科学,2001,29(2):250-252,267.
[2]YC/T311-2009,烤烟品种烘烤特性评价 [S].
[3]藤田茂隆,田岛智之.烤烟易烤性的遗传及香吃味 [J].艾树理,译.中国烟草 ,1984,(3):45-48.
[4]盖钧镒,章元明,王建康.植物数量性状遗传体系 [M].北京:科学出版社,2003.
[5]蔡长春,陈宝元,傅廷栋,等.甘蓝型油菜开花期和光周期敏感性的遗传分析 [J].作物学报,2007,33(2):345-348.
[6]兰 海,余 月,王凤格,等.玉米种子休眠性数量遗传体系的判别 [J].玉米科学,2007,15(2):5-8.
[7]袁有禄,张天真,John Yu,等.棉花高品质纤维性状的主基因与多基因遗传分析[J].遗传学报,2002,29(9):827-834.
[8]陈学军,陈劲枫.辣椒株高遗传分析 [J].西北植物学报,2006,26(7):1342-1345.
[9]韩建明,侯喜林,史公军,等.不结球白菜株高性状主基因+多基因遗传分析[J].南京农业大学学报,2008,31(1):23-26.
[10]王春娥,盖钧镒,傅三雄,等.大豆豆腐和豆乳得率的遗传分析与 QTL定位 [J].中国农业科学,2008,41(5):1274-1282.
[11]王日新,任 民,贾兴华,等.普通烟草栽培种内株高性状主基因加多基因遗传分析 [J].中国烟草科学,2009,30(2):15-20.
[12]王传义.不同烤烟品种烘烤特性研究 [D].北京:中国农业科学院,2008.
[13]Gai JY,Wang JK.Identification and estimation of a QTL model and its effects[J].Theoretical and Applied Genetics,1998,97:1162-1168.
[14]戚存扣 ,盖钧镒,章元明.甘蓝型油菜芥酸含量的主基因+多基因遗传 [J].遗传学报,2001,28(2):182-187.
[15]王传义,张忠锋,徐秀红,等.烟叶烘烤特性研究进展[J].中国烟草科学,2009,30(1):38-41.
[16]Smeeton BW.烟草品质的遗传控制 [J].吴正举,译 .中国烟草,1990,(2):41-48.
