Bt杀虫基因专利保护现状与趋势
2011-06-12束长龙张春鸽
张 杰, 束长龙, 张春鸽
(中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 100193)
在1996-2010年的15年间,全球转基因植物种植面积累计首次超过10亿hm2。2010年全球转基因作物种植面积达到1.48亿hm2,占全球耕地总面积的10%,比1996年增长了87倍,与之相对应的是,2010年转基因作物种子的全球市场价值为112亿美元,转基因玉米、大豆以及棉花产品创造价值约为1500亿美元,这些转基因作物中约39%是具有抗虫性状的,主要是转入了Bt杀虫基因[1]。面对Bt杀虫基因的巨大市场前景,近年来,西方发达国家及其农业生物技术跨国公司不断加大投入,进行新型Bt杀虫基因发掘和相关技术开发,并利用知识产权将这些基因和技术转化为自己独占的权利,以期在未来的竞争中掌控主动权[2-4]。我国于20世纪90年代后期开始了相关的研究工作,与西方发达国家由企业主导的研发不同,我国Bt杀虫基因发掘的主体是国家科研院所与大专院校,目前在国家“863”计划、“转基因生物产业化重大专项”等项目的资助下,呈现出加速发展的良好势态。
本文主要分析Bt杀虫蛋白基因(cry基因)的克隆命名情况,并对目前受专利保护的基因分布状况进行统计,借此为我国相关研究人员在转基因技术研发、产业化应用过程中,合理有效地规避知识产权陷阱,有效利用Bt资源提供决策参考。
1 Bt杀虫蛋白分类
Bt是苏云金芽胞杆菌(Bacillusthuringiensis)的简称,在1901年作为一种昆虫病原菌被发现,从1940年起被大规模开发成微生物杀虫剂,用于害虫防治。Bt特异的杀虫作用一直受到人们的关注,其杀虫物质主要是杀虫晶体蛋白(insecticidalcrystal proteins,简称ICPs),由cry基因和cyt基因编码;随着研究的深入,从Bt中不断分离到新的杀虫活性蛋白,例如营养期杀虫蛋白(vegetativeinsecticidal protein,简称Vips)等[5-7]。
ICPs是Bt主要的毒性物质,具有特异的杀虫活性。1981年,Schnepf首次克隆到cry基因,并证实了其杀虫活性[8]。截止到目前,全球已经克隆并被Bt杀虫毒素国际命名委员会命名的杀虫晶体蛋白基因有 605个,按照命名规则分为72个大类(cry1~cry68,cyt1~cyt4),包括230种模式基因。目前Bt杀虫毒素使用的命名规则是1998年Neil Crickmore博士提出的,杀虫毒素蛋白按照其编码的氨基酸序列相似性进行命名。按照目前的命名规则,所有毒素都可以获得一个包含4个等级的命名(图1)。如果现在有一种新获得的毒素,它与目前所有毒素氨基酸序列相似性都小于45%,属于第1等级的新蛋白,它则会被命名为 Cry69Aa1;与Cry68Aa1的一致性在45%~78%之间则会被命名为Cry68Ba1,属于第2等级的新蛋白;与Cry68Aa1的一致性在 78%~95%之间则会被命名为Cry68Ab1,属于第3等级的新蛋白;而与Cry68Aa1的一致性大于95%则会被命名为Cry68Aa2,属于第4等级的蛋白,以此类推[9]。上述前3类的新毒素与目前已命名毒素序列一致性小于95%,与之相对应的基因通常称为模式基因(holo-typegene),若其杀虫活性高,即可直接申请发明专利。
这些已发现的Cry毒素对无脊椎动物中4个门和节肢动物门中9个目的500余种有害生物具有杀虫活性,并且这些基因拥有各自独特的杀虫谱,如Cry1、Cry3、Cry4、Cry5可分别毒杀鳞翅目、鞘翅目、双翅目等有害昆虫,以及线虫、肝吸虫等;Cry2对鳞翅目、双翅目昆虫有活性;Cry1B、Cry1I对鞘翅目、鳞翅目有活性[10]。目前已有十余种cry基因应用于玉米、棉花、马铃薯和番茄等抗虫作物构建[4]。
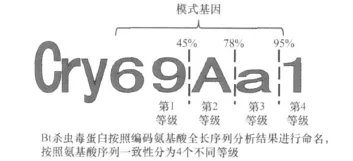
图1 BtCry毒素命名规则
营养期杀虫蛋白(Vips)是苏云金芽胞杆菌在细胞营养期分泌的一种非晶体的胞外杀虫蛋白。Estruch等人于1996年和1998年分别从Bt菌株和蜡状芽胞杆菌(B.cereus)菌株培养物的上清液中分离到Vip杀虫蛋白[5,11]。该类蛋白从对数生长中期开始分泌,直到稳定前期达到最高峰;一般不形成伴胞晶体,且与已知的ICPs序列相似性很低,最重要的是其杀虫作用机理与ICPs不同。该蛋白的发现使Bt在杀虫活性、杀虫谱方面得到很大突破,并在一定程度上克服了多种害虫对ICPs不敏感的弊端。目前Vips命名规则分为4大类(Vip1、Vip2、Vip3和Vip4),其中Vip1与Vip2是一种二元毒素,两者需要协同作用,目前研究显示这类毒素对鞘翅目害虫具有杀虫活性(US5770696,US6656908,US6605701等专利)。Vip3类是一大类对鳞翅目高效的杀虫蛋白,特别是Vip3Aa类蛋白对草地贪夜蛾、小地老虎等害虫有较好杀虫活性,克服了BtCry蛋白对其毒力较低的弱点;Vip4类蛋白是新发现的一类蛋白,其具体功能还不清楚。
除此之外Bt还编码其他杀虫蛋白及其毒力相关的物质,例如几丁质酶、蛋白酶、细菌素、增强素、抗菌肽等,这些物质在Bt杀虫过程中都有重要的作用[12]。此外,Sip1A与S-层蛋白是2006年发现的新型杀虫蛋白[13-14],两者都对叶甲科的害虫有杀虫活性,其杀虫机理尚不清楚。
2 Bt杀虫基因克隆与专利保护现状
2.1 数据来源与分析方法
由于目前应用最广泛的Bt毒素是Cry、Cyt和Vip,本文将主要分析这3种毒素的克隆与专利保护的相关信息。有关基因的克隆与命名主要依据英国萨塞克斯大学(SussexUniversity)生命科学学院Bt毒素命名网站(http:∥www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/)。国外 Bt杀虫基因专利保护情况是通过分析GenBank中专利保护氨基酸(PATAA)数据库获得(http:∥www.ncbi.nlm.nih.gov/genbank/)。PATAA数据库收集了来自美国专利局(USPTO)以及通过欧洲分子生物学实验室(EMBL)、日本DNA数据库(DDBJ)中的欧洲专利、日本专利所涉及的氨基酸序列。分析已经获得正式命名的Bt模式杀虫基因,建立本地杀虫基因数据库,利用本地Bt模式杀虫基因数据库与PATAA数据库中氨基酸序列进行Blastp比对,挑选PATAA数据库中与Bt模式杀虫基因相比得分(Score)在60以上的氨基酸序列进行分析,进一步去除无关序列,获得目前专利保护的所有Bt杀虫蛋白序列。cry基因序列在国内的专利数据,通过对中国专利数据库进行检索,数据库的检索日期是2011年4月1日,包括数据库中此前已公开的所有专利申请文件。采用关键词进行检索,检索词如下:苏云金(杆菌)、苏云金芽胞杆菌、芽胞杆菌、Bt(基因)等,在摘要、说明书和权利要求书中同时检索,并经过人工判读排除不相关专利。
2.2 Bt杀虫基因的克隆与命名
自从1981年第1个杀虫基因被克隆开始,截至2011年4月,有701个杀虫基因被命名,其中cry基因570个,cyt基因35个,vip基因96个,全球Bt基因克隆的速度总体呈现加速上升的趋势(图2)。这些基因来自30多个国家和地区,其中我国最多,有259个。我国Bt基因克隆工作近年来有较大突破,2000年中国农业科学院植物保护研究所克隆到我国第1个模式基因cry1Ie1;2010年我国克隆到的模式基因有31个,占当年全球模式基因总数的89%。

图2 历年我国克隆的Bt基因所占比例
2.3 Bt杀虫基因在国外的专利保护情况
通过对GenBankPATAA数据库氨基酸序列进行比对分析,结果显示目前已授权专利涉及的Bt杀虫基因氨基酸序列有768个,主要分布在对鳞翅目、鞘翅目害虫有活性的 cry1、cry2、cry3、cry8、cry9、cry34和cry35中(图3)。分析结果还显示出一个现象,这些序列中有364条目前已命名的模式基因序列相似性小于95%,按照目前命名规则应属于新型模式基因,占所有受保护序列的47%。这些新基因没有在任何正式的学术刊物上发表,而只是以专利形式出现,这些结果说明国外相关研究机构和企业已经对杀虫基因命名与发表文章不感兴趣,而致力于占有新基因的知识产权。

图3 受专利保护的Bt基因序列类型与数量
近年来,在国家“863”、“转基因专项”等项目的支持下,国内不少研究机构正在开展Bt基因的分离与转基因工作。由于目前国外大量受专利保护的Bt基因并未提交Bt新毒素国际命名委员会命名,序列等信息处于隐藏状态,而这批基因与已知的模式基因的一致性都小于95%。这种情况就给我国许多从事Bt新基因分离与转基因研究的人员带来潜在的知识产权隐患。因为大家很可能不清楚自己所掌握的新型模式基因是否已经被国外专利所覆盖和保护,极易落入知识产权陷阱,进而导致在与国外的知识产权竞争中处于绝对劣势。随着国外大型企业申报中国专利等知识产权申请的逐年增多,对其专利所保护基因信息的准确掌握显得尤为重要。
本文按照目前Bt基因命名规则对已授权专利中与目前已报道的模式基因的一致性小于95%的Bt基因进行分类,统计相关专利号,以供国内从事Bt基因相关研究人员参考。表1列出了与目前已命名的模式基因序列相似性大于78%小于95%的序列相关的专利,按照现行命名规则,应属于第3等级的新模式基因。表2列出了与目前已命名的模式基因序列相似性大于45%小于78%的序列相关的专利,按照现行命名规则,应属于第2等级的新模式基因。表3列出了与目前已命名的模式基因序列相似性小于45%的序列相关的专利,按照现行命名规则,应属于第1等级的新模式基因。
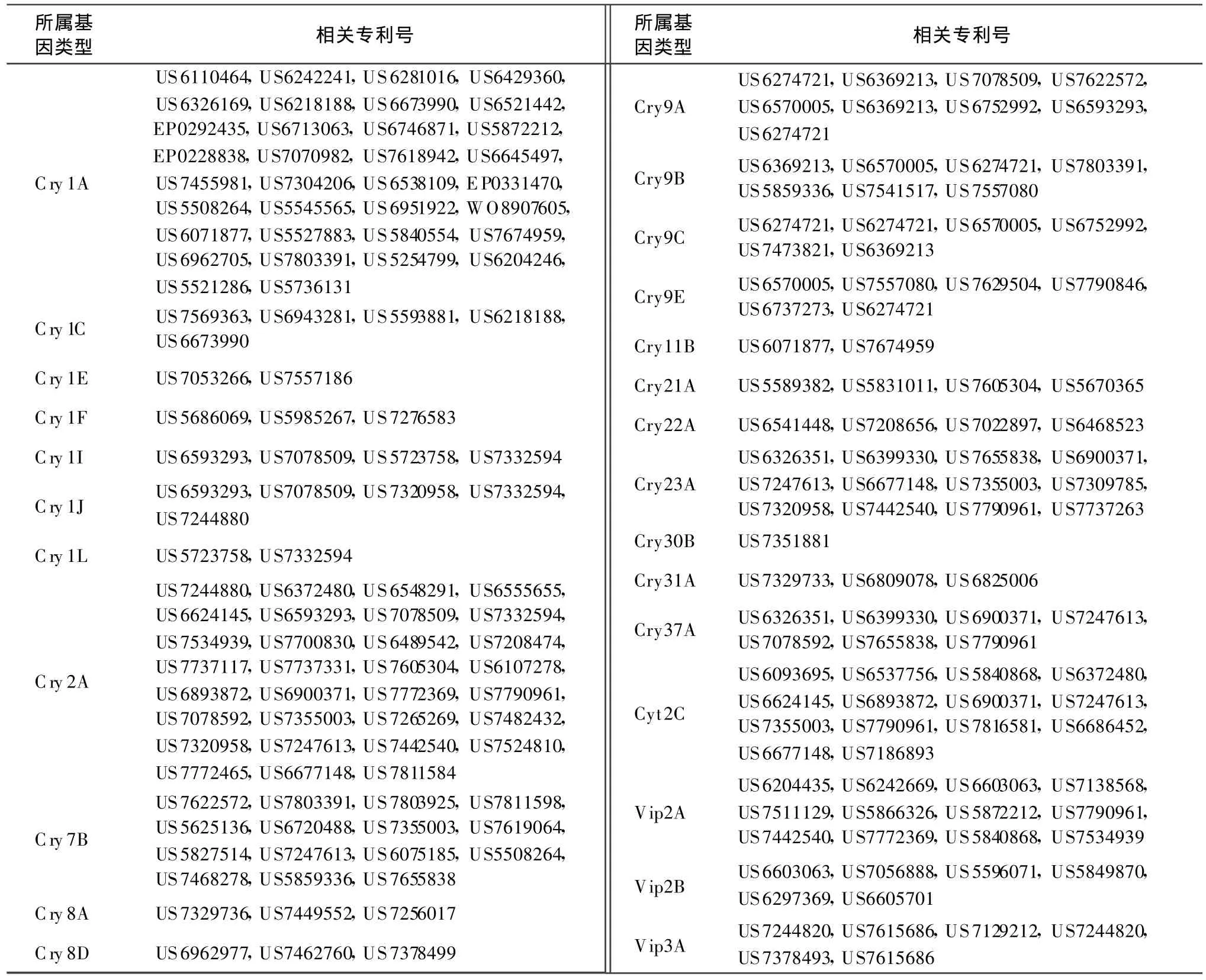
表1 涉及第 3等级的 Bt新基因的专利
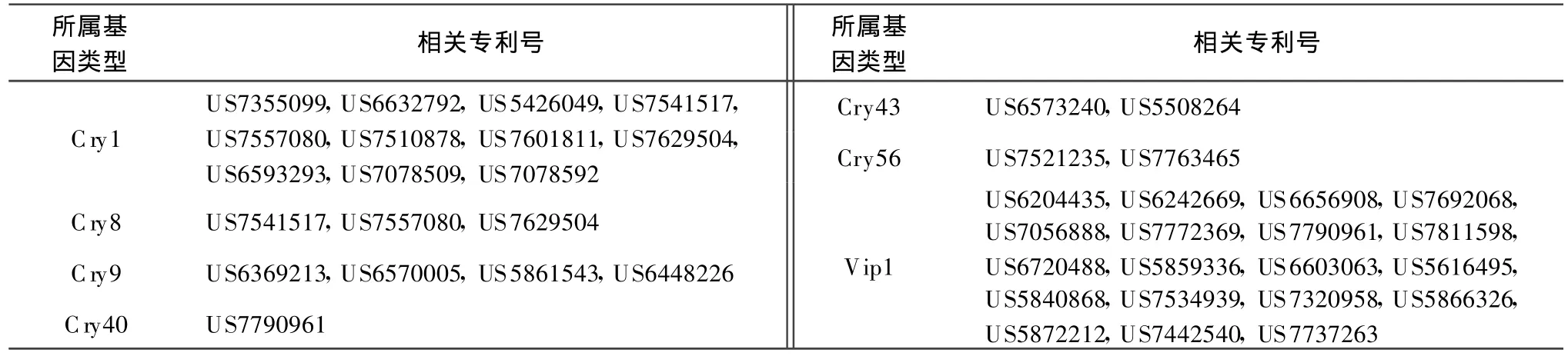
表2 涉及第 2等级的 Bt新基因的专利
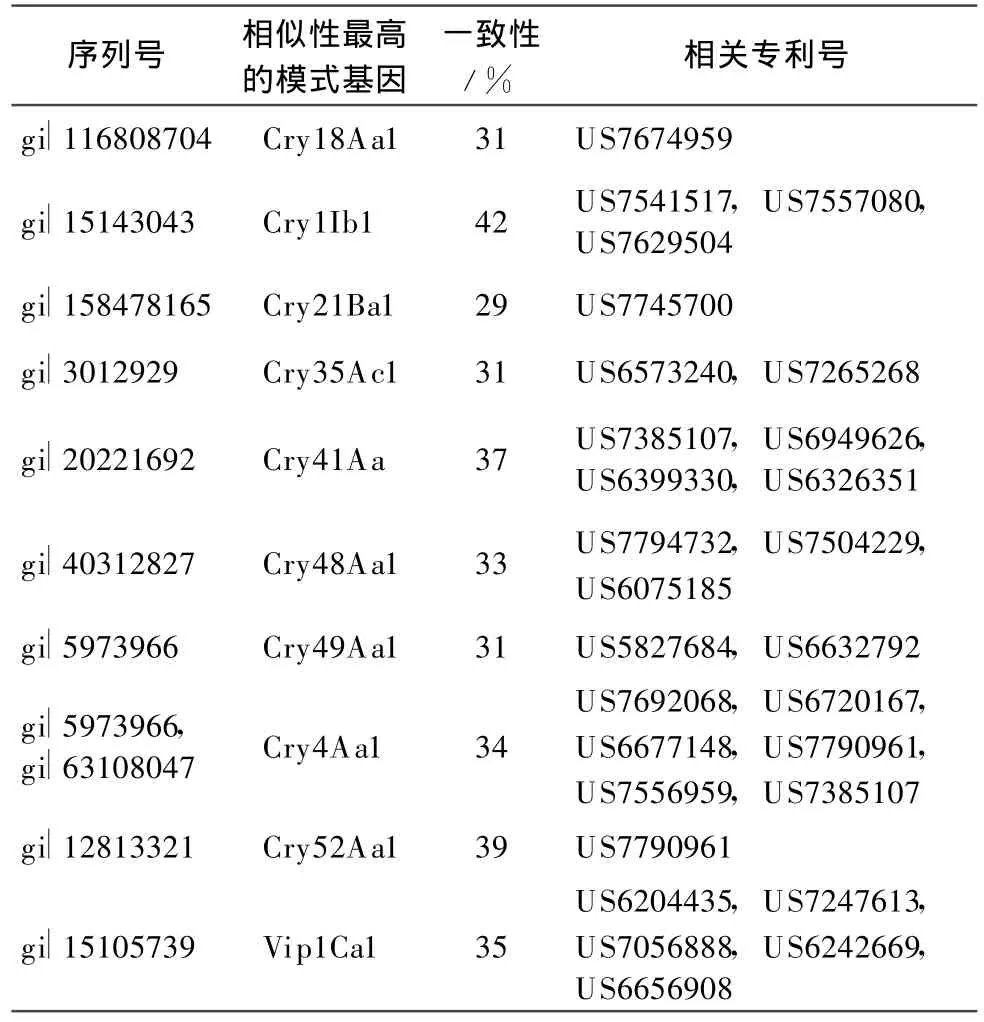
表3 涉及第1等级的Bt新基因及其专利
2.4 Bt基因在国内的专利保护现状
通过检索,获得Bt相关专利260件,剔除Bt菌株、基因改造及Bt杀虫剂相关专利,获得Bt杀虫基因相关专利47件,其中国内研究机构申请的有21件,占44.7%。在受保护基因类型标注清楚程度来看,国内研究机构申请的专利标注明显、准确,通过专利判读,可以准确了解专利所保护的基因类型;而国外研究机构申请的专利对其权利要求基因往往不标注,或者标注不准确,这种现象近年来尤为突出。比如2011年先锋国际良种公司在华申请的5个专利中,在摘要、题名甚至专利全文都没有说明其保护准确基因名称。而2004年拜尔生物科学公司专利(zl1484702)所要求的“cry2Ae” ,“cry2Af” ,“cry2Ag”3 个基因中 ,只有cry2Ae与目前毒素命名网站(http:∥www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/)命名一致,而“cry2Af”、“cry2Ag”则与命名网站对应模式基因序列有较大差异,属于公司个体行为,不能按目前基因命名规则对其准确描述。此外,从申请专利数量上看,国外研究机构加速了在中国知识产权的布局,2010年申请的8件专利中,国内外各占4件,与之前比例相当;而2011年4月份之前申请的6件专利中,国外就占到5件,整体呈现速度加快的趋势。表4列出的是外国研究机构在中国申请专利保护的基因情况,以供相关研究人员参考。
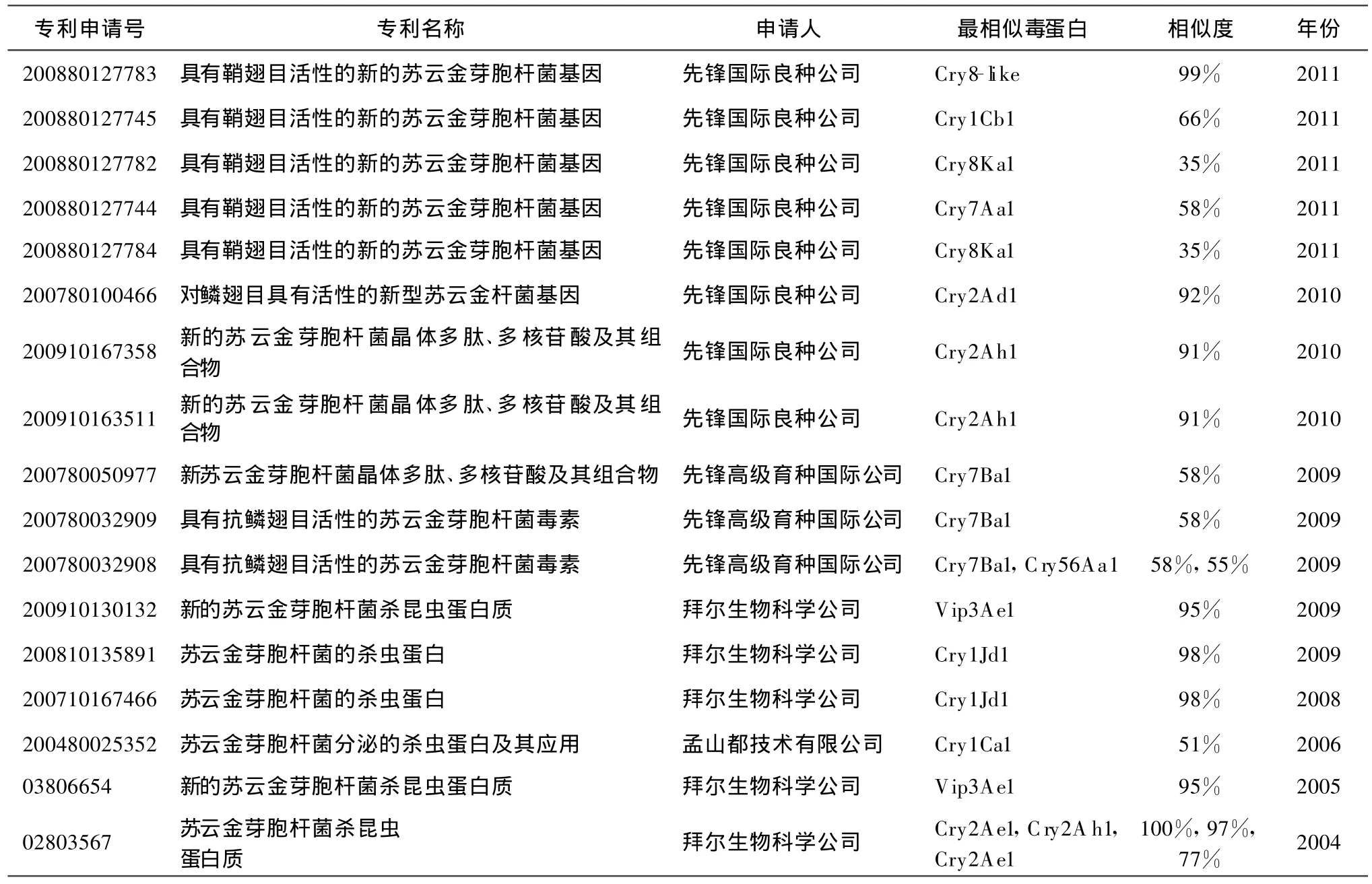
表4 2000-2011年国外企业在中国申请的Bt基因专利

续表4
3 总结与展望
通过对Bt基因命名网站的数据分析统计,结果显示我国相关研究取得了令人瞩目的成就,有大量新型模式基因被分离克隆,特别是近年来所命名的新基因占全球新基因的绝大部分。而当我们分析国外授权专利所保护的Bt杀虫基因的时候发现,虽然国外研究机构命名的Bt基因数目在减少,但是其专利保护的新型Bt基因序列数量却是非常巨大的,并呈上升趋势。这些基因通常不在正式的学术刊物上发表,只有通过检索相关专利数据库才能获得相关信息;即便检索到相关专利,由于其专利保护基因标注不明确,无法获取这些新型基因的更多信息,这些情况容易导致国内相关研究人员在与国际同行竞争过程中陷入被动挨打的境地。
针对这种情况,密切跟踪、整理和分析国外主要农业生物技术研究机构和跨国公司所拥有的Bt基因专利状况是当前一项刻不容缓的工作。国内Bt基因研究人员应当密切跟踪外国主要农业生物技术研究机构的专利申请动态,通过生物信息学、情报学研究方法对其进行整理,形成易读的研究报告,为我国相关管理部门和科研人员提供决策依据和参考。
目前,国内Bt基因专利数量与质量都处于劣势,要改变目前状况需要从多方位进行创新[4]。首先,国外企业在申请专利的推动力方面占有较大优势。获得具有开发前景的Bt基因、并将这些基因和技术转化为独占的权利从而获得最大化的经济利益是国外大型企业的推动力,是其生存与发展的根本。而国内研究主体是科研院所与大专院校,主要依靠国家研究项目推动Bt基因研究,这使得国内研究机构在Bt基因知识产权方面动力不足,难以在竞争中占据优势。目前,主要国外公司掌握了全世界70%以上的Bt杀虫基因专利,并且被批准商业化的cry、vip3A基因转化事件几乎全被孟山都、陶氏益农、先锋、先正达、拜耳等公司掌控。其次,国内单位上中下游脱节,缺乏整体有机的密切配合机制。国外大型企业不仅掌握了绝大部分基因,并且这些基因涉及的专利质量大多高出国内单位的专利,其权利要求范围涵盖较宽、技术全面,包含了基因、载体、宿主菌、转化方法等Bt基因应用的各个方面,从上游至下游技术都囊括其中。这样的专利文件最大限度地保证了其相关基因在应用过程中的生命力,避免了竞争对手在其中某一环节对其基因专利进行绞杀。形成这样的专利需要一个集成了Bt菌株分离与基因克隆平台、农业害虫饲养与测定等功能验证平台、基因改造修饰技术平台、转基因植物技术平台,甚至作物品种资源与选育技术平台的大型研究机构进行全面系统的合作。在国外,孟山都、先锋等公司都具备这样的实力,相比之下国内研究机构和国内企业显得势单力薄,没有一家研究单位或企业可以包揽所有的环节流程,而又缺乏高效的合作机制,从而很难同国外企业竞争。因此,创新上中下游合作机制,让企业成为成果产业化主体,协调基因克隆、遗传转化和转基因新品种培育之间的紧密协作,形成利益共享、风险共担的合作机制,加大Bt基因在各个环节中的知识产权保护力度,让市场成为推动国内Bt基因研究与知识产权保护真正动力,是保障我国的转基因植物产业化早日走上良性发展的必由之路。
[1]James C.Global status of commercialized biotech/GM crops:2010[G].International Service for the Acquisition of Ag ri-biotech Applications,2010,Brief 42.
[2]Shu C,Zhang J.Current patents related to Bacillus thuringiensis insecticidal crystal proteins[J].Recent Pat DNA Gene Seq,2009,3(1):26-28.
[3]邹吉承.中国Bt基因相关知识产权专利保护现状及趋势[J].中国棉花,2007(7):2-5.
[4]宋敏,林祥明,刘丽军.Cry基因家族的专利分布研究[J].生物技术通报,2010(1):1-8.
[5]Schnepf E,Crickmore N,Van R J,et al.Bacillusthuringiensis and its pesticidal crystal proteins[J].Microbiol M ol Biol Rev,1998,62(3):775-806.
[6]Bravo A,Likitvivatanavong S,Gill S,et al.Bacillus thuringiensis:A story of a successful bioinsecticide[J].Insect Biochem Mol Biol,2011(2):1-9.
[7]Mohamed A I,Natalya G,Matthew J,et al.Bacillus thuringiensis A genomics and proteomics perspective[J].BioengineeredBugs,2010,1(1):31-50.
[8]Schnepf H E,Whiteley H R.Cloning and expression of the Bacillus thuringiensis crystal protein gene in Escherichia coli[J].Proc Natl Acad Sci USA,1981,78(5):2893-2897.
[9]Crickmore N,Zeigler D R,Feitelson J,et al.Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins[J].Microbiol Mol Biol Rev,1998,62(3):807-813.
[10]de Maagd R A,Bravo A,Crickmore N.How Bacillus thuringiensis has evolved specific toxins to colonize the insect world[J].Trends Genet,2001,17(4):193-199.
[11]Estruch J J,Warren G W,Mullins M A,et al.Vip3A,a novel Bacillus thuringiensis vegetative insecticidal protein with a wide spectrum of activities against lepidopteran insects[J].Proc Natl Acad Sci USA,1996,93(11):5389-5394.
[12]Raymond B,Johnston P R,Nielsen-Leroux C,et al.Bacillus thuringiensis:an important pathogen?[J].T rends in Microbiology,2010,18(5):189-194.
[13]Donovan W P,Eng leman J T,Donovan J C,et al.Discovery and characterization of Sip1A:A novel secreted protein from Bacillus thuringiensis with activity against coleopteran larvae[J].Appl Environ Microbiol,2006,72(4):713-719.
[14]Pena G,Miranda-Rios J,de La R G,et al.A Bacillus thuringiensis S-layer protein involved in toxicity against Epilachna varivestis(Coleoptera:Coccinellidae)[J].Appl Environ Microbiol,2006,72(1):353-360.
