青藏高原东缘10种禾本科植物种子萌发的基温和积温
2011-04-25王梅英卜海燕
王梅英,刘 文,刘 坤,卜海燕
(1.甘肃省玛曲县草原工作站,甘肃 玛曲 747300;2.兰州大学干旱与草地生态教育部重点实验室,甘肃 兰州730000)
种子萌发连接种子阶段和生长阶段,是完成植物生活史、实现种群更新和物种延续的重要环节[1-2]。温度是影响种子萌发的关键因子,而且对高寒植物的影响尤为重要[3]。温度影响着种子的萌发能力、萌发速率和在萌发时期内的萌发频率[4]。为了描述某个特定物种种子萌发的温度范围,研究者提出了影响种子萌发的3个重要温度:基温(base temperature,Tb),即种子能够萌发的最低温度;最适温度(optimum temperature,To),即萌发速率达到最快时的温度;最高温度(ceiling temperature,Tc),即允许萌发的最高温度[5]。不同物种的这3个重要温度是不同的,与其环境适应范围相关,并与有利于随后幼苗生长的萌发时间的选择相匹配[6]。Garcia-Huidobro等[7]1982年首次提出了积温模型计算种子萌发所需的积温。物种间萌发对温度的不同响应还与物种的地理分布和生态分布有一定的关系[8]。
禾本科植物大约出现在7 000万年前,目前世界上有600~700属,近10 000种,是单子叶植物中仅次于兰科的第二大科。我国共有200余属,1 500种以上。该科植物分布广泛,能够适应各种不同类型的生态环境,既有为人类提供粮食的农作物,也有为家畜提供高营养价值饲料的栽培和野生牧草。禾本科植物也是高寒草甸植被组成的主要组分种和优势种,其种子萌发过程的研究对探讨高寒草甸植被恢复和生态保护有很重要的意义[9-12]。
本研究通过比较分析10种禾本科植物种子萌发所需积温以及基温,试图探讨和解释该区域植物种子萌发过程对青藏高原特殊温度环境的适应对策,为研究高寒草甸物种动态和群落结构提供理论基础。
1 材料与方法
1.1研究地概况 研究区域在甘肃省甘南藏族自治州境内,位于青藏高原东部地区(100°45′~104°45′ E,33°06′~35°70′ N),海拔2 000~4 200 m;年均降水量450~780 mm,主要分布在 7-9月;年平均气温1.8 ℃,最冷月1月平均气温-10.7 ℃,最热月7月平均气温为11.7 ℃,生长季最高气温为23.6~28.9 ℃,年平均霜期不少于270 d,气候特点是高寒湿润。植被以灌木和多年生草本为主[13]。
1.2研究材料 试验选取了禾本科5个属中的10个种(每属选2个种),属分别是羊茅属(Festuca)、鹅观草属(Roegneria)、披碱草属(Elymus)、雀麦属(Bromus)、早熟禾属(Poa)。物种名、生活型和采集地点的海拔见表1。
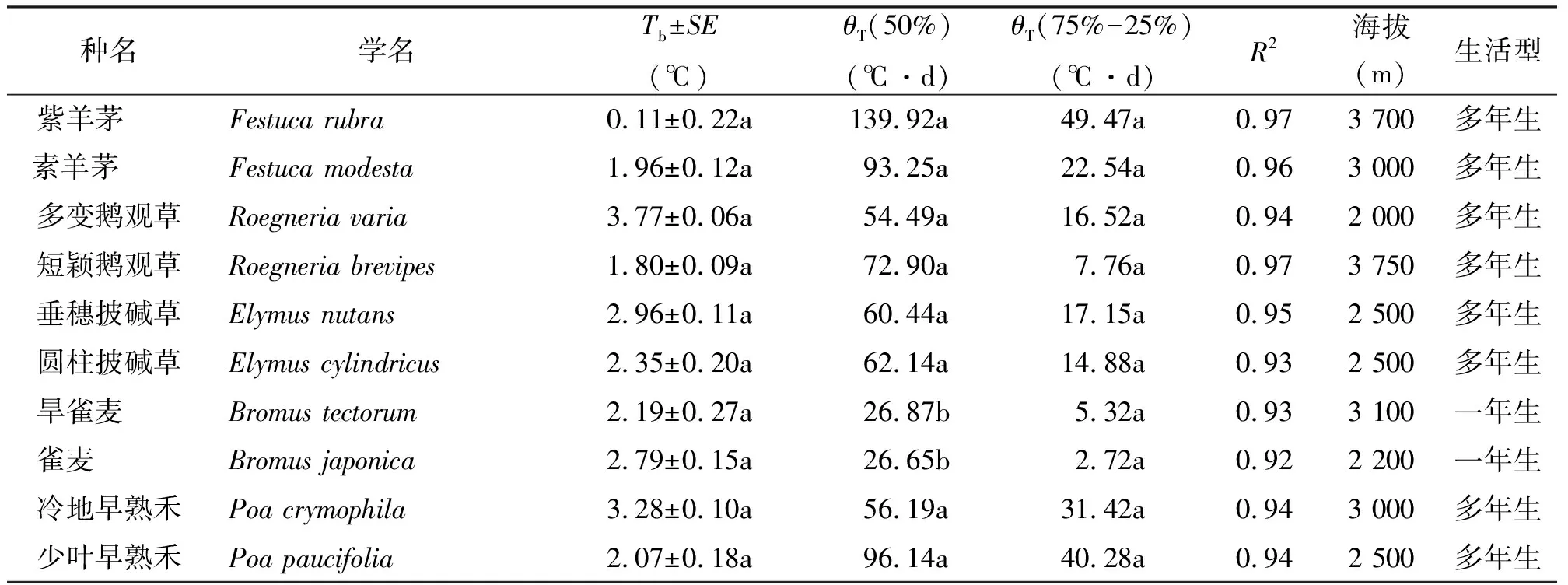
表1 不同植物的海拔、生活型、种子萌发的基温、积温平均值和积温标准差
1.3试验设计 试验所用的种子为2006年6-10月在甘南藏族自治州境内采集。种子均在其自然脱落时收集。所采集的种子清理之后,装在信封内在实验室风干(温度为15~20 ℃)。2007年3月初使用人工气候箱(Conviron E15 growth chamber,Controlled Environments Ltd.Canada)进行萌发试验。试验设置5个温度梯度(5、10、15、20、25 ℃),萌发在黑暗条件下进行。每个种设置3个重复,每个重复随机选取50粒完整种子。将种子均匀放在直径90 mm玻璃培养皿中,培养皿底部垫有两层滤纸,不定期加水保持滤纸湿润。试验过程中每天统计萌发个数,以胚根与种子等长作为萌发的标准,并将已萌发的种子移除,试验持续60 d[12,14]。
1.4研究方法 本试验设置的5个恒温梯度范围(5~25 ℃) 基本上在所研究物种萌发的亚适宜温度范围内,因此,用积温模型来定量分析种子萌发对亚适宜温度的响应。模型假设种子萌发需要累积一定的积温或者热量,同一种群内不同个体的种子萌发速率是不同的[7]。在未达到适宜温度时,萌发速率(用萌发时间的倒数表示)通常是随着萌发温度线性增加的[15]。种子萌发过程中累积萌发率达到g时所需要的积温可以用下面的积温模型公式来计算:
θT(g)=(T-Tb)tg
(1)
式中,T是试验的温度,Tb是萌发的基温,tg是累积萌发率达到g时所需的试验天数。积温是在萌发基温Tb的基础之上累积计算的。在种群内,对所有种子个体来说,Tb通常是固定的值,但积温是不同的,而且是服从正态分布或对数正态分布的[16-17]。
Brown和Mayer[18]用Logistic函数对累积萌发率g和萌发时间t做非线性拟合:
g=m/(1+e-kt+b)
(2)
式中,m近似等于最终萌发率,k是萌发率的增长率,b是与萌发时滞相关的参数。结合公式1和2,累积萌发率g和积温θT(g)也同样满足Logistic函数。因此,可以得到:
g=m/[1+e-k(T-Tb)t+b]
(3)
式中,m是累积萌发率,(T-Tb)t是积温,4个定值参数m,k,b,Tb的值通过非线性回归得到。非线性拟合过程是对累积萌发率在积温水平上完成的。由此通过方程3可以得出达到最终萌发率的g分位时所需要的积温θT(g)的方程:
(4)
用达到最终萌发率的50%所需要的积温θT(g)(50%)和达到最终萌发率的75%和25%所需要的积温的差值θT(75%-25%)分别来近似估计种子萌发所需积温的平均值和标准差[19]。数据处理时将试验所做的3个重复的每日萌发种子数做平均,然后再求得每日的累积萌发率。在做非线性拟合时累积萌发率是因变量,温度和时间是自变量。统计分析和非线性拟合均在R软件 (version 2.10.1)中完成。
2 结果
2.1物种的萌发基温和积温的差异 10个物种萌发的基温范围为0.11~3.77 ℃,积温均值范围为26.65~139.92 ℃·d,积温的标准差范围为2.72~49.47 ℃·d(表1)。羊茅属两个种萌发的积温均值最高,达到93.25和139.92 ℃·d,积温的标准差也较大。紫羊茅在所研究的物种中萌发基温最低,接近于0 ℃。鹅观草属、披碱草属和早熟禾属萌发的基温和积温均值的差异比较小。这3个属物种萌发的基温在2~4 ℃,除了少叶早熟禾萌发的积温均值为96.14 ℃·d之外,其他物种萌发积温均值在50~70 ℃·d。雀麦属中的雀麦和旱雀麦都为一年生的草本,萌发积温均值在26 ℃·d左右,萌发积温的标准差分别为2.72和5.32 ℃·d,积温均值和积温的标准差在5个属中最低。一年生植物种子萌发的积温显著低于多年生植物(P<0.05),但是两者的萌发基温之间和积温的标准差之间差异不显著(P>0.05)。
2.2海拔与萌发基温和积温之间的关系 10个物种的种子采集地点的海拔范围为2 000~3 750 m。对10个物种进行海拔与萌发基温和积温的相关性分析(图1)发现,海拔与萌发基温之间存在显著的负相关性(P<0.05),但是海拔与萌发积温之间的相关性不显著(P>0.05),即随着海拔的升高,物种的萌发基温降低,但是萌发积温无显著变化。

图1 海拔与萌发所需基温和积温之间的关系
3 讨论
青藏高原地带年平均气温低,生长季短,植物为了完成其生活史,需要在春季尽早萌发。因此,适应该地区的植物种子可以在温度较低的条件下萌发。青藏高原的春季,气候复杂多变,如果萌发需要的积温低,植物种子在早春短暂的高温条件下即可迅速萌发,但随之而来的低温就会使其刚出土的幼苗大量死亡。相比之下,如果萌发需要的积温高,植物种子就不会在暂时的高温条件下大量萌发,这样就减小了植物在这样特殊的气候条件下因为暂时的高温萌发所经历的风险。因此,本研究的物种具有萌发基温低和积温高的特点,这是长期适应青藏高原特殊的气候条件的结果。
物种萌发基温的差异在一定程度上反映它们生长环境的差异,这表明物种萌发的基温适应于它们生长的环境[20]。在植物群落建植过程中,萌发和出苗的时间影响物种间的竞争平衡[21-22]。本研究中海拔与萌发基温之间存在显著的负相关性(P<0.05)。一般认为,随着海拔的升高,温度降低。高海拔地区环境恶劣,植物生长季短[23],物种的萌发基温较低,一部分种子在较低的温度条件下能够萌发,幼苗可以尽早进入营养生长,有利于以后在群落中的竞争,这也是对当地气候条件长期适应的结果。本研究表明,海拔对萌发积温的影响不显著,可能是由于物种的萌发积温主要受供试物种的起源和系统发育特征的影响。
Trudgill等[24]的研究表明,一年生和两年生的植物比多年生的植物萌发需要的积温低,然而两者的萌发基温差异不显著。本研究中,雀麦和旱雀麦属于一年生草本,萌发积温也表现出类似的特点。一年生物种的萌发积温较低,萌发持续时间比较短,就能够在适宜温度条件下一次快速萌发,优先占领生存空间,并快速生长完成生命周期,比多年生的物种具有时间和空间上的优势[3,25]。
不休眠种子在一系列温度下的萌发响应与植物的生态地理分布、系统发育和物种不同的生活史对策相关[8,24]。Trudgill等[24]的研究中,生长在英国的羊茅(F.ovina)萌发的基温为6.7 ℃,积温均值为44 ℃·d,紫羊茅萌发的基温为4.7 ℃,积温均值为56 ℃·d,普通早熟禾(P.trivialis)萌发的基温为5.6 ℃,积温均值为43 ℃·d。Larsen和Bibby[19]的研究中,生长在丹麦的两个紫羊茅品种萌发的基温分别为2.7和2.5 ℃,积温均值为60和67.9 ℃·d。与这些研究结果相比,本研究中10种禾本科物种萌发的基温较低,积温较高。
4 结论
本研究中的10种禾本科植物种子萌发的基温较低,积温较高,这是本区域物种长期适应青藏高原特殊气候条件的结果。海拔与萌发基温之间存在显著的负相关性(P<0.05),说明种子萌发的基温与其所处的温度环境相适应。多年生物种的积温较高,一年生物种的萌发所需积温较低。
[1]Moore P D,Chapman S B.Methods in Plant Ecology[M].2nd edition.Oxford:Blackwell Scientific Publications,1986.
[2]樊宝丽,马全林,张德奎,等.露蕊乌头的种子萌发及幼苗出土对不同家系和亲代处理的响应[J].草业科学,2010,27(9):97-103.
[3]徐秀丽,齐威,卜海燕,等.青藏高原高寒草甸40种一年生植物种子的萌发特性研究[J].草业学报,2007,16(3):74-80.
[4]Kocabas Z,Craigon J,Azam-Ali S N.The germination response of Bambara groundnut (Vignasublerrannean(L.) Verdo) to temperature[J].Seed Science and Technology,1999,27:303-313.
[5]Bewley J D,Black M.Seeds:Physiology of Development and Germination[M].2nd edition.New York:Plenum Press,1994.
[6]Alvarado V,Bradford K J.A hydrothermal time model explains the cardinal temperatures for seed germination[J].Plant,Cell and Environment,2002,25:1061-1069.
[7]Garcia-Huidobro J,Monteith J L,Squire G R.Time,temperature and germination of pearl millet (PennisetumtyphoidesS.& H.).I.Constant temperature[J].Journal of Experimental Botany,1982,33:288-296.
[8]Probert R J.The role of temperature in the regulation of seed dormancy and germination[A].In:Fenner M.Seed:The Ecology of Regeneration in Plant Communities[C].2nd edition.Wallingford:CAB International,2000:261-291.
[9]刘振恒,徐秀丽,卜海燕,等.青藏高原东部常见禾本科植物种子大小变异及其与萌发的关系[J].草业科学,2006,23(11):53-57.
[10]王一峰,王春霞,杨文玺,等.甘肃禾本科饲用植物资源研究[J].草业科学,2005,22(4):9-13.
[11]杜玉红,周学丽,王建锋.高寒地区4种禾本科牧草的生产特性比较[J].草业科学,2010,27(8):166-168.
[12]刘明霞.青藏高原高寒草甸禾本科植物种子萌发特性研究[D].兰州:兰州大学,2009.
[13]吴征镒.中国植被[M].北京:科学出版社,1980.
[14]Steadman K J,Pritchard H W.Germination ofAesculushippocastanumseeds following cold-induced dormancy loss can be described in relation to a temperature-dependent reduction in base temperature(Tb)and thermal time[J].New Phytologist,2003,161:415-425.
[15]Bierhuizen J F,Wagenvoort W A.Some aspects of seed germination in vegetables.I.The determination and application of heat sums and minimum temperature for germination[J].Scientia Horticulturae,1974,2:213-219.
[16]Covell S,Ellis R H,Roberts E H,etal.The influence of temperature on seed germination rate in grain legumes.I.A comparison of chickpea,lentil,soyabean and cowpea at constant temperatures[J].Journal of Experimental Botany,1986,37:705-715.
[17]Dahal P,Bradford K J,Jones R A.Effects of priming and endosperm integrity on seed germination rates of tomato genotypes[J].Journal of Experimental Botany,1990,41:1431-1439.
[18]Brown R F,Mayer D G.Representing cumulative germination.2.The use of the Weibull function and other empirically derived curves[J].Annals of Botany,1988,61:127-138.
[19]Larsen S U,Bibby M B.Differences in thermal time requirement for germination of three turfgrass species[J].Crop Science,2005,45:2030-2037.
[20]Trudgill D L.Why do tropical poikilothermic organisms tend to have higher threshold temperatures for development than temperate ones[J].Functional Ecology,1995,9:136-137.
[21]Harper J L.Approaches to the study of plant competition[A].In:Milthorphe F L.Mechanisms in Biological Competition[C].Cambridge:Cambridge University Press,1961:1-39.
[22]Black J N,Wilkinson G N.The role of time of emergence in determining the growth of individual plants in swards of subterranean clover (TrifoliumsubterraneumL.)[J].Journal of Agricultural Research,1963,14:628-638.
[23]郭淑青,齐威,王玉林,等.青藏高原东缘海拔对植物种子大小的影响[J].草业学报,2010,19(1):50-58.
[24]Trudgill D L,Squire G R,Thompson K.A thermal time basis for comparing the germination requirements of some British herbaceous plants[J].New Phytologist,2000,145,107-114.
[25]李巧峡,李凯,赵庆芳,等.不同温度和化学处理对四川嵩草种子萌发的影响[J].西北师范大学学报(自然科学板),2008(5):78-82.
