施肥对玛曲县高寒草甸植物多样性及生产力的影响
2011-04-25牛得草张永超江世高
韩 潼,牛得草,张永超,江世高,傅 华
(兰州大学草地农业科技学院 农业部草地农业生态系统学重点开放实验室,甘肃 兰州 730020)
玛曲县位于青藏高原东端,甘、青、川三省交界地带,是黄河上游最重要的水源涵养地,每年补充黄河水量约占黄河总水量的45%,被誉为“黄河之肾”[1]。然而,近年来由于过度放牧及气候变暖引起的干旱等原因[2-3],该区草地退化、沙化日趋严重,鼠害频繁,草地群落物种多样性减少,生态系统稳定性降低[4-5],生产力不断下降。全县近90%的天然草地出现了不同程度的退化和沙化,其中沙化面积达50 000 hm2以上,且每年以10.8%的速度蔓延[6]。同时高寒草原由于其特殊地理和气候条件,生态系统十分脆弱,极易受到破坏且难以恢复,加之人们更多的看重短期经济效益而忽视长远生态效益,以草原退化为主要标志的生态问题已十分突出[7]。因此,进行退化草地的恢复和治理,对保护草原生态环境、提高牧草产量、增加载畜量和牧民的经济收入具有极其重要的意义。
研究表明[8],采用围栏封育、短时间内利用草地是防止草地退化的有效方法之一。但针对大面积退化草地的恢复重建,最迫切的任务是提高退化草地的土壤肥力,加速牧草繁衍能力和改善牧草的种群结构[9]。草地合理施肥是退化草地恢复重建的关键技术措施之一[10]。草地施肥通过补充土壤营养物质不仅能显著增加草地初级生产力,而且能改善草地植物的营养品质。在高寒草地施肥封育和划破草皮改良措施中,施肥对草地可食牧草产量的增加最为显著[11]。近年来,我国草地施肥研究及推广应用逐渐增多,但其所施肥料主要集中在化学肥料上,主要的研究内容有肥料种类的选择、施肥时期、施肥方法与肥效等方面[12-15]。研究表明,氮、磷、钾及中微量元素肥料合理施用对牧草的产量、品质及对食草动物的生长、发育均会产生明显的促进作用[12],尤其是施用氮肥可以显著增加草地地上生物量,促进禾本科草的生长[12-15]。而根据邱波等[16]研究表明,氮、磷肥混施的效果存在一个阈值,随着施肥量的过量增加,施肥作用会相对减弱。李楠等[17]对草地施肥的研究结果表明,施用尿素、重过磷酸钙等提高了土壤中的碱解氮、硝态氮和速效磷等指标。但在生产实践中,施用化学肥料势必会导致成本增加,尤其在玛曲地区化肥的使用还将会污染黄河水域生态系统。因此,该地区采用厩肥还田,既可避免使用化肥成本高、污染环境等问题,又可充分利用当地现有资源,变肥为宝,达到改良退化草地的目的。然而,关于施畜肥对于退化高寒草甸草地生态系统影响的研究不多[18],本研究探讨了短时间内植物群落对不同施肥梯度下的响应特征,以期为高寒草甸生态系统的恢复提供一定的技术与理论依据。
1 材料与方法
1.1试验地概况 研究区位于青藏高原东部的甘肃省玛曲县阿孜畜牧科技园区,地处33°24′17″ N,101°31′59″ E,海拔3 585 m。属于高原大陆性气候,年均温度1.2 ℃,1月平均温度-10 ℃,7月平均温度11.7 ℃。降水主要集中于5-9月,年降水量为564 mm,年蒸发量1 000~1 500 mm。全年日照时数2 613.9 h,全年霜期大于270 d,没有绝对的无霜期[18]。土壤类型为亚高山草甸土。植物类型以莎草科的嵩草属(Kobresia)和禾本科的针茅属(Stipa)、羊茅属(Festuca)、早熟禾属(Poa)的一些种,以及菊科的风毛菊属(Saussurea)、橐吾属(Ligularia)和瑞香科的狼毒属(Stellera)等毒杂草为主,并伴有其他杂类草,如高山唐松草(Thalictrumalpinum)、莓叶委陵菜(Potentillafragarioides)、钝裂银莲花(Anemoneobtusiloba)、细叶亚菊(Ajaniatenuifolia)、火绒草(Leontopodiumhastioides)、花苜蓿(Medicagoruthenica)等[19]。
1.2试验设计 在研究区选择退化程度均一、地势平坦的退化草地,于2010年5月进行围栏封育。试验设置4个处理:不施肥为对照(S0);施有机肥11.25 t/hm2(S1);施有机肥22.50 t/hm2(S2);施有机肥33.75 t/hm2(S3),所施肥料为羊粪。每个处理设5个重复小区,每个小区面积为15 m×10 m,进行随机区组排列。待土壤解冻后,于2010年5月中旬将有机肥撒施到各小区,之后统一进行划破处理,以提高施肥效果。
1.3植被调查 2010年8月中旬,在不同处理的小区内随机设置1个面积为0.5 m×0.5 m的样方,分别进行群落学调查,记录每个样方内的植物种类、密度、株高、盖度。然后,将样方内的植物齐地面剪下,分种装进信封,带回实验室置于烘箱内,105 ℃杀青30 min,然后在65 ℃温度下烘24 h后称质量,以计算植物生物量。群落植物分为3个功能群:莎草类、禾草类、杂类草[20]。
1.4数据处理
1.4.1植物群落多样性分析的计算
Margalef丰富度指数(R)=(S-1)/lnN;
Simpson 多样性指数(D)=1-∑Pi2;
Shannon-Wiener多样性指数(H)=∑PilnPi;
Pielou均匀度指数(E)=H/lnS。
式中,S为每个样地样方中的总物种数,N为每个样方中的全部物种的总个体数,Pi为样方中第i种的生物量占全部物种生物量的比例。
1.4.2统计分析 采用Excel进行制图,SPSS 13.0进行数据统计分析,单因子方差分析(ANOVA)和最小显著差数法(LSD)用于不同施肥强度下各指标间的比较和差异显著性检验(P<0.05)。
2 结果与分析
2.1不同施肥梯度下群落盖度与高度分析 施肥后,S1、S3处理的植物群落盖度同对照(S0)存在显著差异(P<0.05),但各施肥处理(S1、S2和S3)间差异不显著(P>0.05)(图1)。各施肥处理(S1、S2和S3)间的植物群落高度与对照(S0)差异不显著(P>0.05),施肥处理S1同S3之间存在显著差异(P<0.05)(图2)。
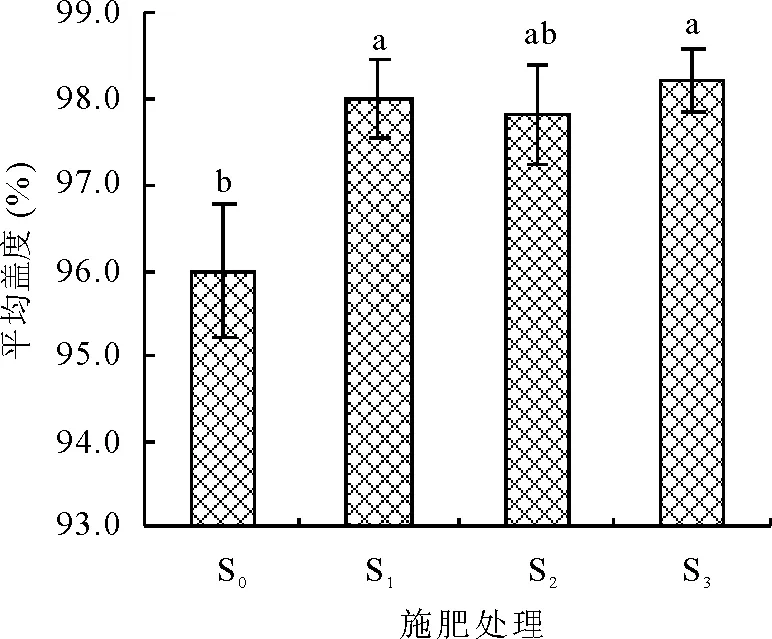
图1 植物群落平均盖度对施肥强度的响应
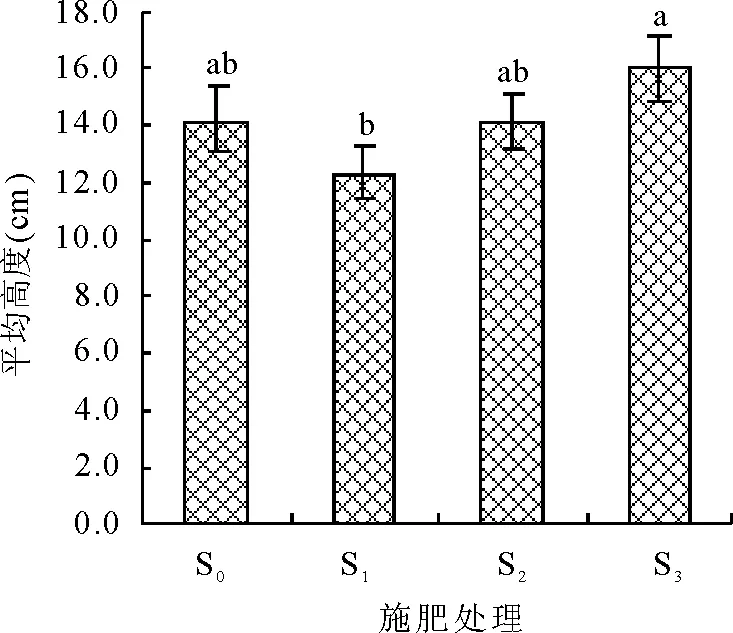
图2 植物群落平均高度对施肥强度的响应
2.2不同施肥梯度下植物群落地上总生物量及各功能群生物量的变化 各施肥处理(S1、S2和S3)的植物群落地上总生物量同对照(S0)相比,尽管差异不显著(P>0.05),但是都表现出明显的增加趋势,分别较对照增加了9.6%、17.8%和30.9%(图3)。施肥后,各施肥处理(S1、S2和S3)的莎草类生物量显著高于对照(P<0.05),但各施肥处理(S1、S2和S3)间差异不显著(P>0.05),禾草和杂类草生物量较对照无显著差异(P>0.05)。
植物群落功能群的相对生物量在不同施肥处理间表现明显变化(图 4)。施肥处理显著提高了群落中莎草类的比例(P<0.05),但各施肥处理(S1、S2和S3)间差异不显著(P>0.05);禾草类和杂类草比例有所减少,但较对照差异不显著(P>0.05)。
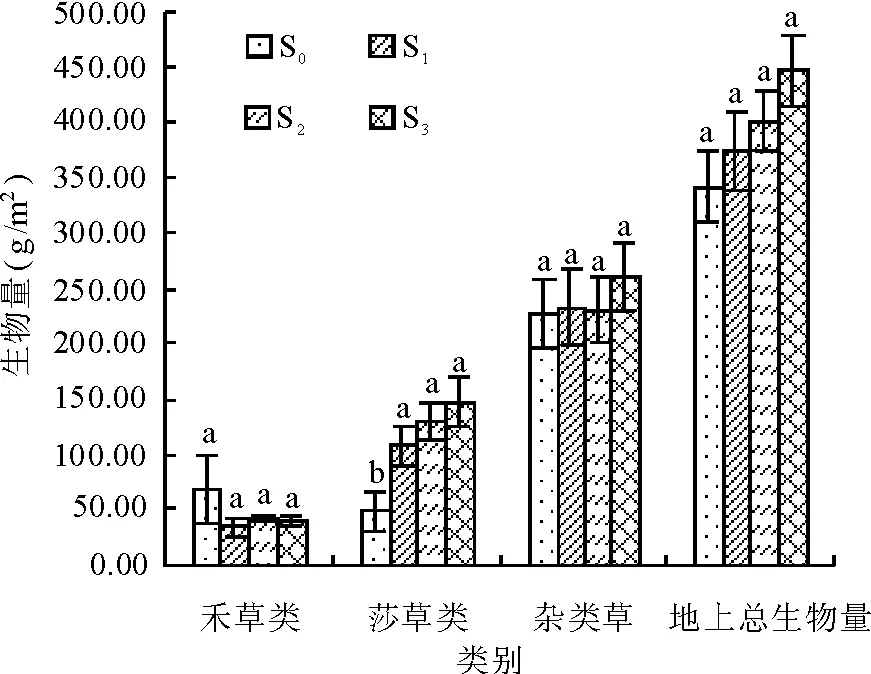
图3 植物功能群生物量与地上总生物量对不同梯度施肥处理的响应
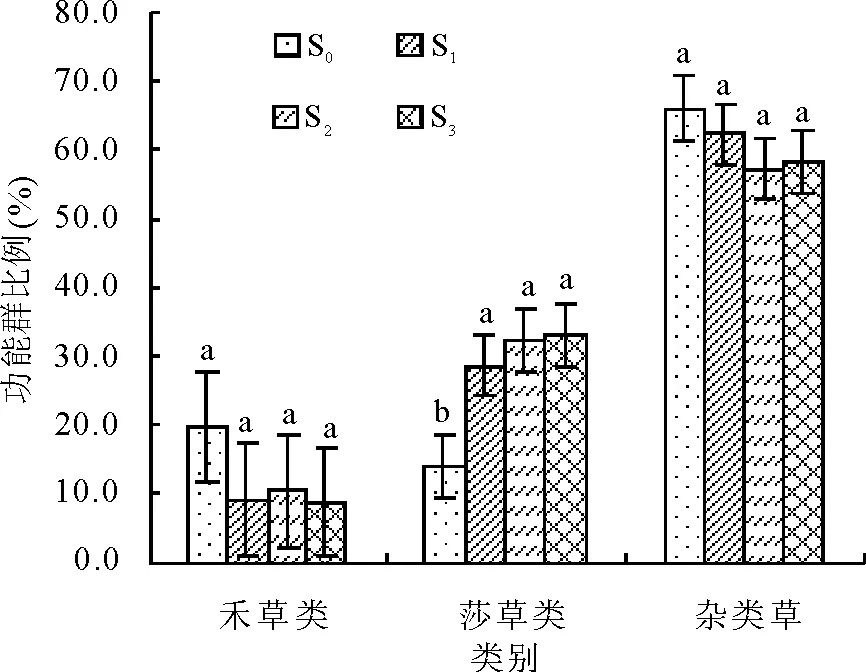
图4 植物功能群生物量所占比例对不同梯度施肥处理的响应
2.3不同施肥梯度下植物群落物种多样性变化 施肥后,群落丰富度指数变化显著(表1),各施肥处理(S1、S2和S3)的物种数与Margalef指数均高于对照(S0)且差异显著(P<0.05),而对于群落的均匀度指数而言,对照(S0)明显高于各施肥处理(S1、S2和S3),但无显著差异(P>0.05)。Shannon-Wiener物种多样性指数变化不明显,而Simpson物种多样性指数则表现为S1和S2施肥处理下低于对照(S0),施肥处理S2、S3与对照(S0)处理差异不显著(P>0.05)。
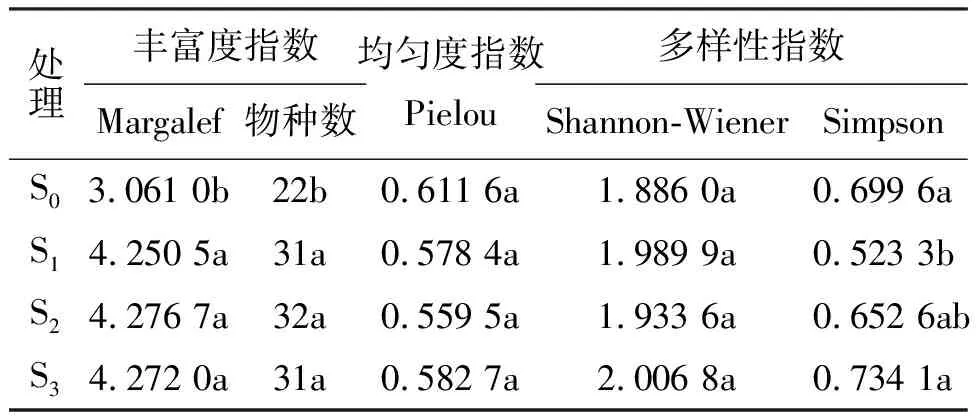
表1 植物群落中丰富度指数、多样性指数和均匀度指数对施肥强度变化的响应
3 讨论
草地施肥后其群落高度和盖度往往显著提高,沈景林等[15]研究发现草地施肥7年后植被盖度分别较对照增加18.7%~71.2%。邱波等[16]研究发现草地施肥3年后其群落植被高度显著提高。本研究也表明,在施肥当年,施肥处理就使得群落盖度大于对照,尤其施有机肥11.25 t/hm2处理与施有机肥33.75 t/hm2处理同对照相比达显著水平,但是,施肥后群落植被高度与对照相比差异不显著。这可能是由于施肥处理时间较短,施肥对群落高度造成的效应还未表现出来。因为施肥作为一种改变植物群落的干扰手段,其主要是通过改变土壤的理化性状来间接影响群落中植物种空间分布与组成特征的,所以施肥后植物群落结构显著变化很可能需要一个较长的过程。
施肥后一般都会使得植物群落总生物量增加,而各功能类群的生物量的响应却不尽相同。韩文武和杜国祯[21]的研究表明,施肥处理显著增加了草地的地上总生物量,另外,草地施肥后草地禾草生物量与豆科、莎草科、毒杂草类等功能群生物量的变化呈负相关关系。纪亚君[12]也报道,草地施肥后,植物群落禾草类生物量逐渐减少。本研究草地施肥后,尽管草地地上总生物量各处理之间没有显著差异,但随着施肥量的增加表现出明显增加的趋势,这可能是由于有机肥的施用对土壤营养成分具有良好的改良作用。根据刘振恒等[10]的研究结果,每公顷施加15 000~22 500 kg羊粪则相当于每公顷施加氮肥135.0~202.5 kg,磷肥60~90 kg,钾肥105.0~157.5 kg。对于植物群落而言,氮素通常为其主要的营养限制因素[22]。杜国祯和王刚[23]的研究结果表明氮与磷均是高寒草甸植物群落的限制因子。这也说明了施用有机肥料对于高寒草甸植物群落的恢复具有较好的作用。伴随总生物量的变化,群落各功能群植物生物量及在群落中的比例往往也发生改变。群落中属于同一功能群类型的不同植物,其往往是作为一个相对统一的整体对生态因子波动或对外界干扰做出反应的[24]。本研究表明,施肥处理后草地莎草类生物量及花群落中的比例显著高于对照,而禾草和杂类草生物量较对照无显著差异,不过两者在群落中比例有减少的趋势。此结果一定程度上反映了高寒草甸草地植物群落结构上的正向演替模式,即以杂草为主的草甸群落→莎禾草与杂草共存的草甸群落→以莎禾草为主的草甸群落[25]。
群落多样性指数受物种丰富度指数和均匀度指数两个方面的共同影响[26]。均匀度是群落中不同物种的多度分布的均匀程度,即组成群落种群的配置状况,一般认为群落的均匀度越高,群落的稳定性,群落物种多样性也就越高[27]。群落的均匀度较小,导致多样性指数变小。陈亚明等[19]的研究结果表明,施肥后的Shannon-Wiener指数则维持在1.347 4~3.156 9之间,而Margalef指数和Pielou指数则会降低。本研究中草地施肥后群落的均匀度同对照相比减小,多样性指数亦位于上述范围之内;物种数与Margalef丰富度指数同对照相比却明显增加且呈显著差异,出现该种现象的原因尚不明确,需要进一步探讨。
[1]张志强,孙成权,吴新年,等.论甘南高原生态建设与可持续发展战略[J].草业科学,2000,17(5):59-64.
[2]Wang W Y,Wang Q J,Wang H C.The effect of land management on plant community composition,species diversity,and productivity of alpineKobersiasteppe meadow[J].Ecological Research,2006,21(2):181-187.
[3]Wang G,Qian J,Cheng G,etal.Eco-environmental degradetion and causal analysis in the source region of the Yellow River[J].Environmental Ecology,2001,40:884-890.
[4]Zhou H K,Zhou L,Zhao X Q,etal.Stability of alpine meadow ecosystem on the Qinghai-Tibetan Plateau[J].Chinese Science Bulletin,2006,51(3):320-327.
[5]May R M.Stability and Complexity in Model Ecosystems,Population Biology Monographs[M].New Jersey:Princeton University Press,1973:21-38.
[6]尚燚,秦建宏,尚小生,等.玛曲县草地畜牧业存在的问题及可持续发展对策[J].青海草业,2006,15(2):28-30.
[7]戎郁萍,韩建国,王培.不同草地恢复方式对新麦草草地土壤和植被的影响[J].草业学报,2002,11(1):17-23.
[8]周国英,陈桂琛,赵以莲,等.施肥和围栏封育对青海湖地区高寒草原影响的比较研究Ⅰ群落结构及其物种多样性[J].草业学报,2004,13(1):26-31.
[9]李本银,汪金舫,赵世杰,等.施肥对退化草地土壤肥力、牧草群落结构及生物量的影响[J].中国草地,2004,26(1):14-17.
[10]刘振恒,陈听,梁国栋,等.甘南玛曲草地退化现状及恢复重建初探[J].草业与畜牧,2006(8):21-24,29.
[11]Shen J L,Meng Y,Hu W L.Experimental study of improment measures on the degradated grassland in alpine areas[J].Acta Pralaculturae Sinica,1999,8(1):9-14.
[12]纪亚君.青海高寒草地施肥的研究概况[J].草业科学,2002,19(5):14-18.
[13]袁宏波,王辉,李晓兵,等.玛曲县天然草地沙化动态及现状分析[J].甘肃农业大学学报,2006,41(1):73-78.
[14]车敦仁.青海高寒牧区禾草施磷施氮的增产效应[J].草业科学,1990,7(5):15-20.
[15]沈景林,谭刚,乔海龙,等.草地改良对高寒退化草地植被影响的研究[J].中国草地,2000.22(5):49-54.
[16]邱波,罗燕江,杜国祯.施肥梯度对甘南高寒草旬植被特征的影响[J].草业学报,2004,13(6):65-68.
[17]李楠,龚长虹,刘伟,等.施肥对草原土壤养分动态变化的影响[J].安徽农业科学,2009(1):88-89.
[18]张云,武高林,任国华.封育后补播“高寒1号”生态草对玛曲退化高寒草甸生产力的影响[J].草业科学,2009,26(7):99-104.
[19]陈亚明,李自珍,杜国祯.施肥梯度对高寒草甸植物多样性和经济类群的影响[J].西北植物学报,2004,34(3):424-429.
[20]刘蓉,张卫国,江小雷,等.垂穗披碱草群落退化演替的植被特性及其与土壤性状的相关性研究[J].草业科学,2010,27(10):96-103.
[21]韩文武,杜国祯.青藏高原高寒草甸植物群落退化过程和恢复机理的研究[D].兰州:兰州大学,2006.
[22]Bryan L F,Katherine L.Cross species richness in a successional grassland: Effects of nitrogen and plant litter[J].Ecology,1998,79(8):2593-2603.
[23]杜国祯,王刚.甘南亚高山草甸人工草地的演替与质量变化[J].植物学报,1995,37(4):306-313.
[24]邢福,祝廷成,李宪长.松嫩平原羊草草地植物功能群组成及多样性特征对水淹梯度的响应[J].植物生态学报,2002,26(6):708-716.
[25]武高林,任国华.放牧强度对青藏高原东部高寒草甸植物群落特征的影响[J].草业学报,2009,18(5):256-261.
[26]Gao X M,Ma K P,Huang J H,etal.Study on plant community in Dong lingshan Mountain,Beijing,China:Ⅺ.The diversity of mountain meadow[J].Acta Ecology Sinica,1998,18(1):24-32.
[27]Wang S H P,Li Y H,Wang Y F,etal.Influence of different stocking rates on plant diversity ofArtemisiafrigidacommunity in Inner Mongolia steppe[J].ACTA Botanica Sinica,2001,43(1):89-96.
