青海草地早熟禾栽培草地植被特征及土壤物理性状动态
2011-04-25杨慧茹马玉寿李世雄王彦龙伊晨刚
杨慧茹,马玉寿,2,李世雄,2,盛 丽,2,王彦龙,2,伊晨刚
(1.青海大学,青海 西宁 810016;2.青海省畜牧兽医科学院,青海 西宁 810016)
生态学理论认为各种各样的生物和非生物因子决定着陆地植物群落的结构和动态。在空间和时间尺度上,气候变化、人类干扰、环境的严酷性、环境异质性和生物间的相互作用决定着植物群落组成和多样性[1]。在草地生态系统中植被与土壤之间构成一个相互作用、相互影响、相互制约、协调发展的统一体。植被的演替和退化会引起土壤特征的改变,而土壤的变化同样会引起植被的改变[2]。为保护青藏高原的生态环境,恢复治理退化草地需要充分了解栽培草地演替过程中的一些生物学和土壤学过程和特征。青海草地早熟禾(Poapratensiscv.Qinghai)是近年来驯化选育出的牧草新品种,由于它具有发达的根茎,营养繁殖能力极强,能快速形成草皮,是三江源区“黑土滩”植被恢复的适宜草种之一。但是目前关于青海草地早熟禾的研究主要集中于高产农艺措施[3]、建植栽培草地恢复“黑土型”退化草地[4-5]以及生理生化[6]等方面,而对青海草地早熟禾栽培草地植被特征、土壤物理性状变化的研究相对较少。为此,从不同建植年限青海草地早熟禾栽培草地群落特征和土壤物理性状变化入手,探讨高寒地区退化草地恢复过程中植被和土壤的变化过程,旨在为草地生态系统的恢复和栽培草地的合理利用及适应性管理提供理论依据。
1 材料与方法
1.1研究区概况 研究在青海省果洛藏族自治州玛沁县大武镇格姆滩进行,地理位置32°3l′~35°37′ N,96°54′~101°51′ E,平均海拔3 700 m。属典型高原大陆性气侯,年平均气温-3.9 ℃,≥5 ℃年积温850.3 ℃·d,最冷月(1月)平均气温-12.6 ℃,最热月(7月)平均气温9.7 ℃。牧草生长季为156 d,全年无绝对无霜期。年降水量513.2~542.9 mm,多集中在6-9月,年蒸发量为2 471.6 mm。青海草地早熟禾栽培草地建植之前,试验区是典型的“黑土型”退化草地,草地植被属高寒草甸,原生植被盖度不足10%,牧用价值已基本消失。土壤为高山草甸土。自2001年起,对该类草地通过完全耕翻建立青海草地早熟禾多年生栽培草地,耕翻深度20 cm左右,耙平后条播,行距15 cm,播量7.5 kg/hm,同时施尿素(氮含量46%)75 kg/hm2,过磷酸钙(P2O5含量40%)150 kg/hm2。播种时间为5月下旬。
1.2研究方法
1.2.1野外调查与取样 选择生长年限分别为2年、4年、5年、9年和10年的青海草地早熟禾栽培草地为试验样地。2010年8月中旬在上述5个样地内分别随机选取5个1 m×1 m样方,进行植物群落特征调查,调查内容包括群落总盖度、物种种类及其分盖度(用估测法)、株高(每种植物随机测定10株的自然高度,取平均数)、频度,然后齐地面分种剪草,烘干称量。
1.2.2土壤物理性状测定 2010年8月,选取生长2年和9年的青海草地早熟禾栽培草地,在剪去植物地上部分的样方中测定栽培草地的土壤含水量和土壤容重。土壤含水量采用烘干称量法测定,用土钻分别钻取0~10、10~20、20~30 cm层土样,5次重复,称取鲜质量后在105 ℃烘干至恒质量,然后称取烘干质量,计算出土壤含水量,本研究中土壤含水量是指土壤水质量与干土质量的百分数。用环刀法测定土壤容重。
1.3数据分析 物种多样性采用物种丰富度(S)、多样性指数(Shannon-Wiener index)和均匀度指数(Pielou index)计算。物种丰富度指数为每个样地的物种数,多样性指数和均匀度指数计算公式为:
Pielou指数:J=H′/lnS。
式中,Pi为种i的相对重要值[(相对高度+相对盖度+相对频度)/3];S为种i所在样方的物种总数。
以上数据采用Excel 2003 进行初步整理,用SPSS 18.0 对数据进行方差分析,差异显著性在0.05水平上进行比较,数据以“平均值±标准误”表示。
2 结果与分析
2.1不同建植年限栽培草地多样性特征 植物群落的物种丰富度指一个群落所含的植物种数,稳定的栽培草地群落是物种组成恒定、产量稳定的群落,其演替是杂类草不断侵入、物种丰富度不断升高的过程[7]。青海草地早熟禾栽培草地群落的物种丰富度指数变化范围为12.6~13.8(表1),且不同生长年限栽培草地之间差异不显著(P>0.05)。

草地生态系统的可持续性和草地生产力的维持在很大程度上依赖于草地群落的生物多样性,安渊等[8]研究认为,特定资源生产力水平下草地群落固有的生物多样性是保持草地稳定和健康发展的基础。王国宏[9]认为,群落的稳定性受制于群落物种的多样性。对不同建植年限栽培草地群落多样性测度表明,不同建植年限栽培草地之间Shannon-Wiener指数差异不显著(P>0.05),Pielou指数差异显著(P<0.05),排序为:5年>9年>10年>2年>4年。这与王长庭等[10]对三江源地区不同建植年限栽培草地植被群落多样性变化的研究结果一致。
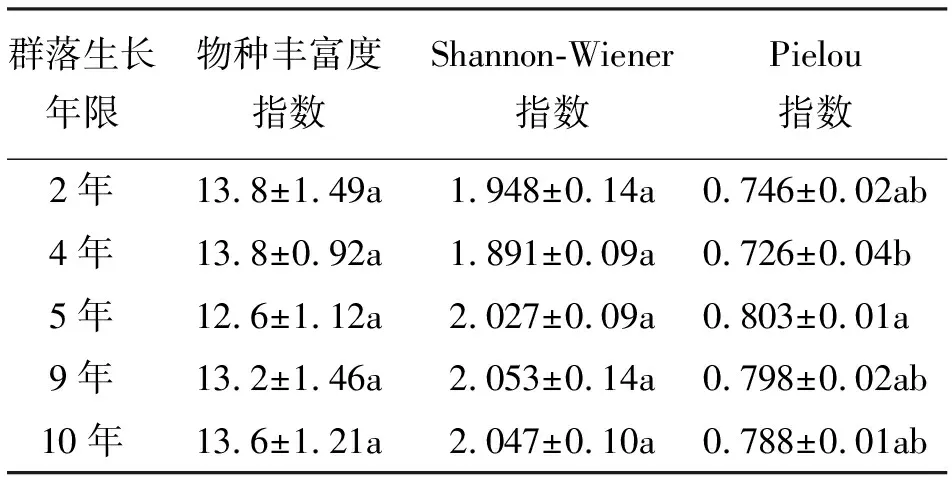
表1 不同生长年限青海草地早熟禾栽培草地物种丰富度指数和多样性指数特征
2.2不同建植年限栽培草地群落结构特征 通过对不同建植年限青海草地早熟禾栽培草地群落结构特征的调查分析表明(表2),群落的总盖度在生长第2年达到98.0%,且与4年、5年、9年和10年栽培草地之间差异显著(P<0.05),随着生长年限的延长,到第10年栽培草地群落总盖度虽然有所下降,但仍然能保持在90%以上。优势种青海草地早熟禾的盖度随着生长年限的延长而降低,且除9年和10年栽培草地之间差异不显著外,其余不同年龄栽培草地之间差异显著(P<0.05)。说明栽培草地建植初期,群落优势种青海草地早熟禾的盖度高,维持着群落的主要生态功能,同时伴随着对严酷环境抵御能力强的一、二年生植物侵入,但随着生长年限的延长,多年生植物替代一、二年生植物,其他物种和杂类草的侵入,导致青海草地早熟禾的盖度不断下降。青海草地早熟禾高度的变化为建植第2年较高,平均达到了71.6 cm,到第4年有所下降,且与第2年差异显著(P<0.05),到第5年植株高度相比第4年有所回升,与第2年和第4年差异显著(P<0.05),随生长年限延长到第9年和第10年,青海草地早熟禾的高度下降速度较慢。不同青海草地早熟禾栽培草地群落中,随着生长年限的延长,植物量呈下降的趋势。通过方差分析可知,2年和4年草地之间地上植物量差异显著(P<0.05),且均与5年、9年和10年栽培草地差异显著,但5年、9年和10年栽培草地之间差异不显著。这种情况可能与不同年间的气候条件(尤其是降水、积温的差异)以及植物群落的演替阶段有关。

表2 不同建植年限青海草地早熟禾栽培草地植被特征
2.3不同建植期栽培草地土壤物理性状 土壤是植物生长的基质,其理化特性决定着植物群落类型的分布,同时植物群落又反作用于土壤,逐步改善其生境条件,使群落得以发展[11]。土壤含水量和土壤容重是表征土壤结构状况和水源涵养功能的物理性质指标。生长2年和9年青海草地早熟禾栽培草地各土壤层含水量差异不显著(P>0.05),但9年栽培草地各土壤层含水量均比2年草地有所上升,这说明随生长年限的延长,青海草地早熟禾栽培草地涵养水源的能力渐渐增强。土壤容重是土壤紧实度的敏感性指标,也是表征土壤质量的一个重要参数,可反映土壤结构、透气性、透水性能以及保水能力的高低。土壤容重越小,说明土壤结构、透气和透水性能越好[10,12-13]。土壤容重在第2年较低,而在第9年相对较高(表3),且二者间除0~10 cm土层之间差异显著(P<0.05)外,其余2层之间差异均不显著(P>0.05),且9年比2年草地各土层的土壤容重增加0.03~0.33 g/cm3,另外随着土壤深度的增加土壤容重逐渐增大。这是因为随着青海草地早熟禾栽培草地生长年限的延长,0~30 cm土层根系多而致密,草地的紧实度逐渐增加,土壤生态系统基本处于相对稳定而良性循环状态。
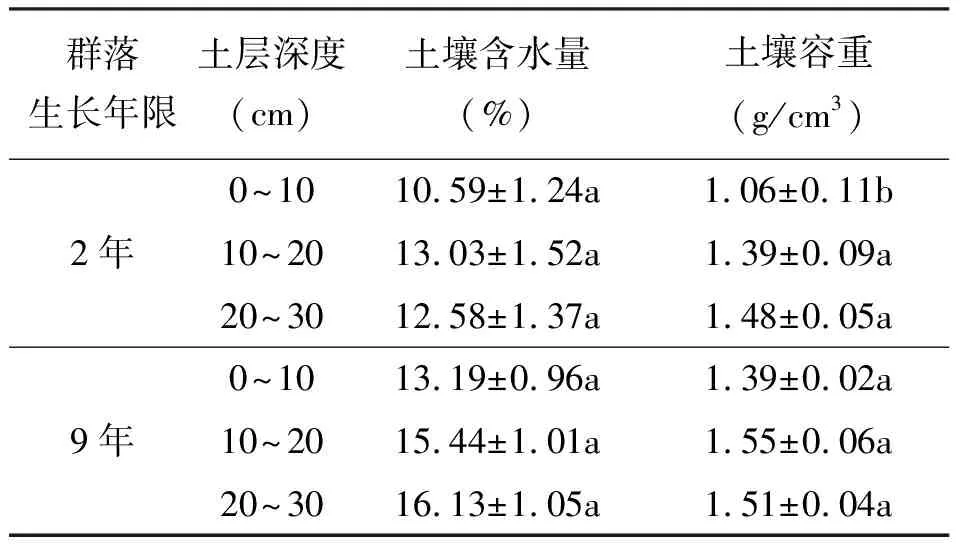
表3 不同生长年限青海草地早熟禾栽培草地土壤物理性状
3 讨论
3.1植物群落的变化 植物群落的演替是对其初始状态的异化过程,不但体现在种类组成和结构上,也体现在土壤环境的改变上。植被与土壤的关系是植物生态学研究的一个重要内容,在同一气候条件下,土壤分异导致植被的变化,植物的生长又可以改善土壤生境[10,14]。本研究中,栽培草地建植第2年,群落总盖度、优势种青海草地早熟禾盖度、植株高度和地上植物量均表现最好,处于生产力最高阶段;到第4年草地群落处于物种组成、生产力比较稳定的阶段,群落优势种青海草地早熟禾维持群落的主要生态功能。植物群落演替主要表现为不同物种的相互替代而导致植物群落在组成结构和功能等方面发生变化。随着利用年限的增加,由于邻近草地的影响和各种植物种子的入侵,栽培草地的物种组成和群落结构逐渐发生改变。建植5年后的栽培草地由于其群落结构发生较大的变化,毒杂草比例增加,群落中原来的优势种青海草地早熟禾减少,青海冷地早熟禾和垂穗披碱草等物种侵入,物种组成明显改变,群落多样性指数逐渐增大,草地牧用价值开始下降;建植9年和10年的草地群落盖度、植株高度和地上植物量较低,但随着杂类草和其他物种的入侵,物种组成逐渐增加,多样性指数有所下降,群落的数量特征虽很低,但就草地植被恢复角度来说,群落逐步趋于稳定。
3.2土壤物理性状的变化 土壤作为植被演替的主要驱动力,其理化特性随栽培草地的建植和群落的演替而改变,土壤环境的变化又影响植物的种间竞争和物种更替,为植物群落的演替奠定了基础[15]。土壤水分是土壤基本组成成分之一,且在土壤物质交换、植物营养等方面起着重要的作用。草地在长期的保护利用下,增强了土壤的吸水和保水能力,缩小了土壤的干层分布深度[16-18]。Martin和Chambers[19]的研究表明,土壤水分含量是河滩草甸植物多样性、植被演替、草地恢复的决定因素,而且土壤通气状况和土壤表面硬度是植物根系生长、光合作用的主要因素。研究结果表明,生长2年和9年的青海草地早熟禾栽培草地土壤含水量、土壤容重有所差别。生长2年和9年的草地,各土壤层含水量差异不显著(P>0.05),但9年栽培草地各土壤层含水量均比2年栽培草地高,这说明随生长年限的延长,青海草地早熟禾栽培草地涵养水源的能力渐渐增强。土壤容重在第2年较低,而在第9年相对较高,且9年比2年草地各土层土壤容重增加0.03~0.33 g/cm3,另外随着土壤深度的增加,土壤容重逐渐增大。这是因为随着青海草地早熟禾栽培草地生长年限的延长,0~30 cm土层根系多而致密,草地的紧实度逐渐增加,土壤生态系统基本处于相对稳定而良性循环状态。土壤物理特性的改变为后来的植物定居创造了有利条件,根系和枯枝落叶回归土壤为微生物的活动提供了能源,群落水热环境和土壤质量的改变使先锋植物在竞争中失去了优势而让位于后来者,植物种类替代加速,从而促进植物群落生物种类多样性和结构复杂性,土壤中物质的输入和输出速率加快,促进了土壤物质循环,使土壤环境得到进一步改善。某一演替阶段土壤的肥力状况,不仅反应了在此之前植被与土壤协同作用的结果,同时也决定了后续演替过程的土壤肥力基础和初始状态。因此,植物群落演替过程,也是植物对土壤不断适应和改造的过程,土壤性质是植物演替的重要驱动力之一[10,20]。
4 结论
不同生长年限青海草地早熟禾栽培草地群落物种数、多样性指数和植被特征都不尽相同,且存在明显差异。对不同生长年限栽培草地群落物种数和多样性测定表明,不同阶段植物群落物种数目上没有显著差异,但在种类组成上却相差甚远;不同生长年限栽培草地之间Shannon-Wiener指数没有显著差异(P>0.05),Pielou指数间差异显著(P<0.05),排序为5年>9年>10年>2年>4年。随着生长年限的延长,青海草地早熟禾栽培草地的群落总盖度、优势种盖度、植株高度和地上植物量都呈下降的趋势,且不同建植年限之间存在显著差异(P<0.05)。此后随着演替进行,其他物种和杂类草的侵入,到第9年和10年草地植被群落特征逐渐趋于稳定。
[1]王长庭,龙瑞军,王根绪,等.高寒草甸群落地表植被特征与土壤理化性状、土壤微生物之间的相关性研究[J].草业学报,2010,19(6):25-34.
[2]董全民,赵新全,李青云,等.小嵩草高寒草甸土壤营养因子及水分含量对牦牛放牧率的响应.Ⅰ夏季草场土壤营养因子及水分含量的变化[J].西北植物学报, 2004,24(12):2228-2236.
[3]王柳英,马玉寿,施建军,等.草地早熟禾高产农艺措施数学模型与优化指标的研究[J].种子,2005,24(1):11-13.
[4]文金花,马玉寿,施建军,等.利用草地早熟禾改建江河源区“黑土型”退化草地的研究[J].草原与草坪,2006(2):41-44.
[5]马金祥,桑永燕,尕松代吉.四种禾草补播“黑土型”退化草地效果初步观测[J].青海草业,2006,15(3):5-6.
[6]文金花,马玉寿,施建军,等.三江源区四种栽培禾草抗寒性研究[J].青海畜牧兽医杂志,2006,36(3):3-5.
[7]王刚,蒋文兰.人工草地种群生态学研究[M ].兰州:甘肃科学技术出版社,1998:232-235.
[8]安渊,李博,杨持,等.内蒙古大针茅草原草地生产力及其可持续利用研究.Ⅰ放牧系统植物地上现存量动态研究[J].草业学报,2001,10(2):22-27.
[9]王国宏.再论生物多样性与生态系统的稳定性[J].生物多样性,2002,10(1):126-134.
[10]王长庭,曹广民,王启兰,等.三江源区不同建植期人工草地植被特征及其与土壤特征的关系[J].应用生态学报,2007,18(11):2426-2431.
[11]Maria B V,Nilda M A,Norman P.Soil degradation related to overgrazing in the semi-arid southern Caldenal area of Argentina[J].Soil Science,2001,166(7):441-452.
[12]董全民,赵新全,马玉寿,等.江河源区披碱草和星星草混播草地土壤物理性状对牦牛放牧强度的响应[J].草业科学,2005,22(6):65-70.
[13]郭艳玲,韩建国,赵守强,等.种植多年生禾本科牧草对土壤物理性状的影响[J].草业科学,2006,23(10):14-18.
[14]Yao H,He Z,Wilson M J,etal.Microbial biomass and community structure in a sequence of soils with increasing fertility and changing land use[J].Microbial Ecology,2000,40:223-237.
[15]王启兰,王长庭,刘伟,等.三江源区不同建植年限人工草地植物群落与土壤微生物生理类群的变化[J].应用生态学报,2009,20(11):2646-2651.
[16]程积民,万惠娥,雍绍萍,等.干旱环境胁迫下草地水分动态与生产潜力[J].草业科学,2002,19(增刊):341-344.
[17]姚拓,马丽萍,张德罡.我国草地土壤微生物生态研究进展及浅评[J].草业科学,2005,22(11):1-7.
[18]田永明,刘德福.农牧交错区草地资源可持续利用指标体系研究[J].草业科学,2004,21(增刊):57-63.
[19]Martin D,Chambers J.Restoration of riparian meadows degraded by livestock grazing: above and belowground responses[J].Plant Ecology,2002,163:77-91.
[20]曲国辉,郭继勋.松嫩平原不同演替阶段植物群落和土壤特征的关系[J].草业学报,2003,12(1):18-22.
