宝天曼自然保护区锐齿栎种群空间分布格局研究
2011-04-23梅世秀袁志良叶永忠朱学灵张志铭刘晓静
梅世秀,袁志良,叶永忠,朱学灵,,张志铭,3,黄 萍,刘晓静
(1.河南农业大学生命科学学院,河南郑州450002;2.国家林业局宝天曼森林生态定位研究站,河南内乡474350;3.中山大学生命科学学院,广东 广州510275;4.河南农业职业学院,河南 中牟451450)
空间分布格局是指生物个体在与环境相互作用中所形成的,种群所在生境内个体的空间配置[1],是种群自身的生物学特性与外界因素综合作用的产物,研究优势种群的分布格局及动态规律,对阐明群落稳定、演替及恢复机理有重要意义[2,3].关于物种空间格局的研究,国内外做了一些研究,研究者从植物种间关系等方面对空间分布格局进行了分析,大多集中于木本植物方面,也有一些方法研究,比如张金屯[4]引入点格局分析,马克明等[5]在将分形理论运用于格局研究等方面做了一些工作.锐齿栎(Quercus aliena var.Acutesrrata)群落是宝天曼自然保护区分布较为广泛的植物群落类型之一,也是该保护区落叶阔叶林的主要成分.目前对该保护区锐齿栎种群空间分布格局方面的研究较少.因此,对锐齿栎种群空间分布格局进行研究,将有助于人们了解锐齿栎种群空间分布状况,了解该种群的稳定状态,为锐齿栎天然林的管理保护、开发利用及营造等方面提供理论依据.
1 研究地区概况
宝天曼自然保护区位于河南省西南部内乡县境内,地处秦岭东段,伏牛山南坡.地理坐标为111.47″~112.04″,北纬33.20″~33.36″,属季风型大陆性气候,四季分明,年均温15.2℃,极端最高温41.2℃,极端最低温-14.8℃,≥10℃的年积温4 200~4 900℃,年降雨量885.6 mm,年蒸发量991.6 mm,相对湿度68%.保护区内主要岩石是花岗岩、石灰岩和砂岩.土壤可划分为山地棕壤、山地黄棕壤和山地褐土3种土类,土壤pH值为6.5左右.
保护区地处暖温带向北亚热带的过渡区,具有丰富的生物多样性,森林覆盖率高达到89%,共有高等植物223科、1 002属、2 711种,分别占河南省高等植物科的87%、属的74%、种的57%.植被以落叶阔叶林、针阔叶混交林为主,主要树种有栎类、鹅耳枥类、槭树类和华山松、油松等[6].
2 研究方法
2.1 样地调查
在宝天曼自然保护区内经充分踏查后,在海拔高度为1 332 m和1 407 m的栎林主要分布区各选取1 hm2连续地段作为调查样地,分别在1 hm2地内设置20 m×20 m的样方25个,再在每个20 m×20 m样方内采用相邻格子法设置16个5 m×5 m的小样方,1 hm2样地内总计有400个小样方.在每个小样方内调查锐齿栎株数并测量胸径和树高.
2.2 数据分析方法
测定生物种群空间分布格局类型的数学模型较多,在实际应用中,各模型均有其适应性和优缺点.为避免不同模型的片面性,本研究采用多个指标进行测度.以样地内锐齿栎定位数据为依据,分析种群的空间分布格局时采用了以下方法:
①离散分布的理论拟合[7].泊松(Poisson)分布用于描述种群的随机分布,其特征是种群中的个体占据空间任一点的概率是相等的,并且任何一个个体的存在不影响其它个体的存在.负二项(negative binomal)分布是描述种群集团分布的一种,其特点是种群在空间的分布呈不均匀的镶嵌状图.为了准确了解种群的分布格局是服从Poisson分布还是服从负二项分布,用X2检验来判断它们的拟合程度.
②方差均值比率[7](v/m):v/m=S2/¯x中,S2为样方内物种个体数的方差,¯x表示为样方内物种平均个体数.当S2/¯x=1时,种群趋于随机分布;S2/¯x>1时,种群趋于聚集分布;S2/¯x<1时,种群趋于均匀分布.该值的显著性可用t检验来确定.
③ 负二项参数[7](K):值用于度量聚集程度,其中K值愈小,聚集程度愈高;当K值趋于无穷大时(一般为8以上),则逼近Poisson分布;K<0时,为均匀分布;K>0时,为聚集分布.
④Cassie指标[4](CA):CA=1/K,式中,K为负二项参数;当CA=0时,为随机分布;CA>0时,为聚集分布;CA<0时,为均匀分布.
⑤ 聚块性指数[4](m*/m)式中m为平均密度,K为负二项参数;当m*/m=1时,为随机分布;m*/m<1时,为均匀分布;m*/m>1时,为聚集分布.
⑥Green指数[7](GI):GI当GI<0时,为均匀分布;GI=0时,为随机分布;GI>0时,为聚集分布.
⑦ 扩 散 型 指 数[7](Iδ):Iδ= N -当Iδ=1时,为随机分布;Iδ>1时,为聚集分布.
⑧ 丛生指数[8](I):-1当I<0时,为均匀分布;I=0时,为随机分布;I>0时,为聚集分布.
以上格式中,¯x为样方内物种平均个体数,S2为样方内物种个体数的方差.
根据特定的时间调查方法,用不同径级大小的个体群来表示不同发育阶段的种群.具体的方法是根据锐齿栎的胸径大小分布的特点及其与生长发育的关系,将锐齿栎种群分为3类个体群:①大树,胸径≥22.5 cm;② 中径树,胸径为7.5 ~22.5 cm;③幼树,胸径<7.5 cm.把野外取样数据根据不同个体群进行处理,然后用上述方法分析不同发育阶段种群的空间分布格局[9].
3 结果与分析
3.1 不同取样尺度下宝天曼自然保护区锐齿栎种群空间分布格局
将海拔梯度为1 407 m样地调查数据应用上述方法进行种群分布格局拟合及聚集强度的测定(表1).由表1可见,在 5 m ×5 m,5 m ×10 m,10 m×10 m,10 m ×20 m,20 m ×20 m 取样尺度上,空间分布格局离散拟合结果均符合负二项分布;方差/均值比率法测定结果均符合聚集分布;在聚集强度测定方面:在以上5个取样尺度上,负二项参数(K)均大于0且小于8,Cassie指标(CA)均大于0,聚块性指数(m*/m)均大于1,丛生指数(I)均大于0,Green指数(GI)均大于0,扩散型指数(Iδ)均大于1,上述这些指标都表明,在5种取样尺度上,锐齿栎种群均符合聚集分布类型,而且不同的取样尺度,种群的聚集程度不同.锐齿栎种群分布格局在不同取样尺度上存在差异,种群在大尺度上明显呈团块状,而在样地尺度上呈现明显的聚集性分布,这种分布格局可能是异质性环境和种内竞争所导致的.在某种意义上讲,它与环境条件的相关是因果关系,或者说种群的空间分布格局特征是种群对环境长期适应的结果,因而种群格局通常反映着一定环境因子对个体行为、生存和生长的影响[8].
在种群分布格局研究中,都具有较大的尺度依赖性,不同的观测尺度所得的结果可能截然不同.这种尺度的依赖性增加了种群分布类型判断的复杂性和不确定性[10].前人研究表明,对于个体呈随机分布的种群,样方面积的变化只会改变样方平均值的大小,而当个体呈聚集分布时,样方大小有可能对结果产生显著影响,如果样方远大于种群分布斑块的大小,测定结果会是一个均匀分布的格局,样方面积太小,取样资料会得出一个随机分布格局[11].
3.2 不同发育阶段宝天曼自然保护区锐齿栎种群的空间分布格局
将海拔梯度为1 407 m样地调查数据,应用上述方法对不同发育阶段锐齿栎种群进行分布格局拟合及聚集强度的测定(表2).由表2可见,在幼树、中径树阶段空间分布格局离散拟合结果均符合负二项分布,而大树符合随机分布;方差/均值比率法测定结果表明幼树、中径树阶段空间分布格局均符合聚集分布,而大树符合随机分布;在聚集强度测定方面:在幼树,中径树阶段,负二项参数(K)均大于0且小于8,Cassie指标(CA)均大于0,聚块性指数(m*/m)均大于1,丛生指数(I)均大于0,Green指数(GI)均大于0,扩散型指数(Iδ)均大于1,上述这些指标都表明,幼树和中经树阶段均符合聚集分布,但聚集程度不同,幼树的聚集程度比中经树高.在大树阶段,扩散系数(C)=1.016,负二项参数(K)=44.531>8,Cassie指标(CA)=0.023,聚块性指数(m*/m)=1.023,丛生指数(I)=0.016,Green 指数(GI)=0.000,扩散型指数(Iδ)=1.068,这些指标都表明大树符合随机分布类型.种群在不同的发育阶段、不同的年龄段会表现出不同的空间分布格局.分析种群在不同发育阶段的空间分布格局变化可以推断种群新生个体的产生、幼树和中径树的生长、大树的死亡以及种群遭受人为干扰的概况[3].目前,乔木种群的分布格局中有关异龄种群的研究较多[12],一般情况下(环境均一,生存条件良好),以母株为中心的繁殖方式使低龄树呈现聚集分布;随着年龄的增加,树木对光因子和营养条件等的要求加大及自疏、它疏等作用,种群会趋于均匀分布;随着年龄继续增加,种群将趋于随机分布[13].但由于物种的不同和生境的差异,这种趋势会发生变化.如戚继忠等研究各类生境中的榛子(Corylus heterophylla)种群均趋于集群分布,随着年龄的增加聚集程度由幼年期的强弱相同,至中年的逐渐减弱,最后过渡到随机分布型[14];王得祥等研究的华山松(Pinus armandi)种群的分布格局随种群发育阶段而变化,总的趋势是由集群分布-随机分布-集群分布[15];刘仁林等研究了南方红豆杉(Taxus mairei)在不同时期种群分布格局,在侵入期表现为随机分布,适应期趋于聚集分布,发展期呈随机分布,稳定期和衰退期趋于聚集分布[16].
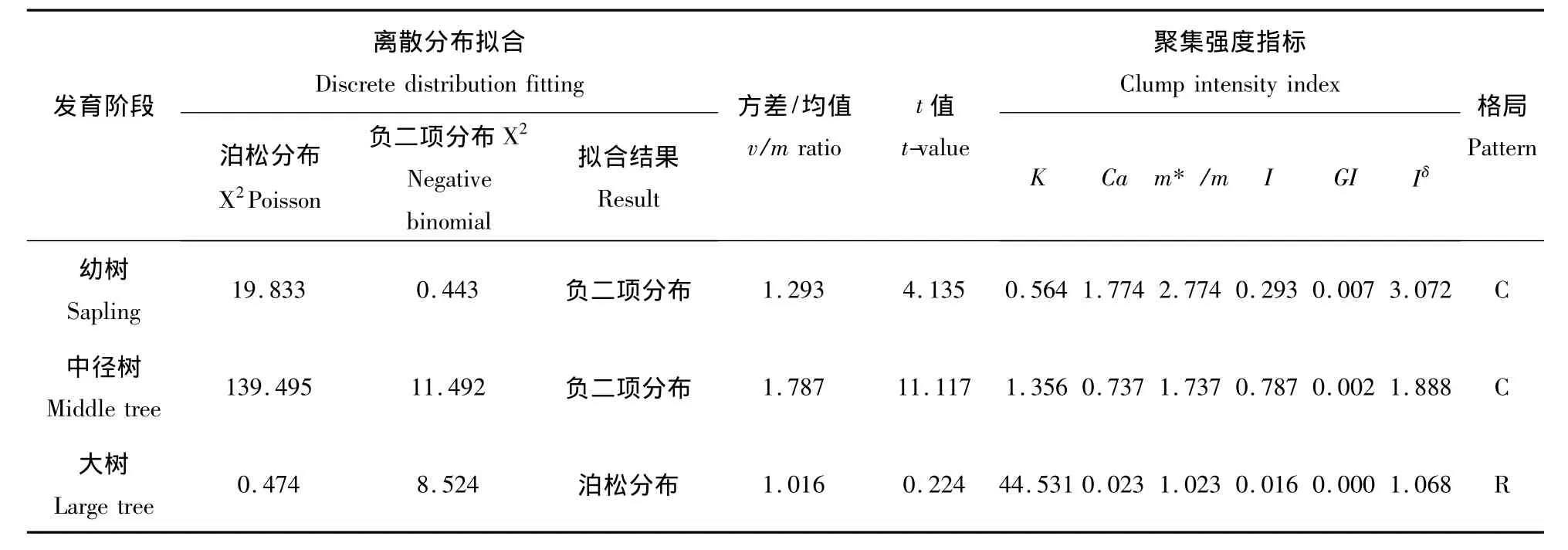
表2 不同发育阶段宝天曼自然保护区锐齿栎种群的空间分布格局Table2 Spatial distribution pattern of Quercus aliena var.Acutesrrata population in Baotianman Nature Reserve at different development stages
3.3 不同海拔梯度宝天曼自然保护区锐齿栎种群的空间分布格局
将海拔梯度为1 332 m和1 407 m样地调查数据,应用上述方法对不同海拔梯度锐齿栎种群进行分布格局拟合及聚集强度的测定(表3).由表3可见,海拔梯度为1 407 m和1 332 m时,空间分布格局离散拟合结果均符合负二项分布;方差/均值比率法测定结果均符合聚集分布;在聚集强度测定方面:负二项参数(K)均大于0且小于8,Cassie指标(CA)均大于0,聚块性指数(m*/m)均大于1,丛生指数(I)均大于0,Green指数(GI)均大于0,扩散型指数(Iδ)均大于1,上述这些指标都表明,海拔高度在1 332 m和1 407 m地区的锐齿栎种群空间分布格局均表现为聚集分布,但聚集程度不同.
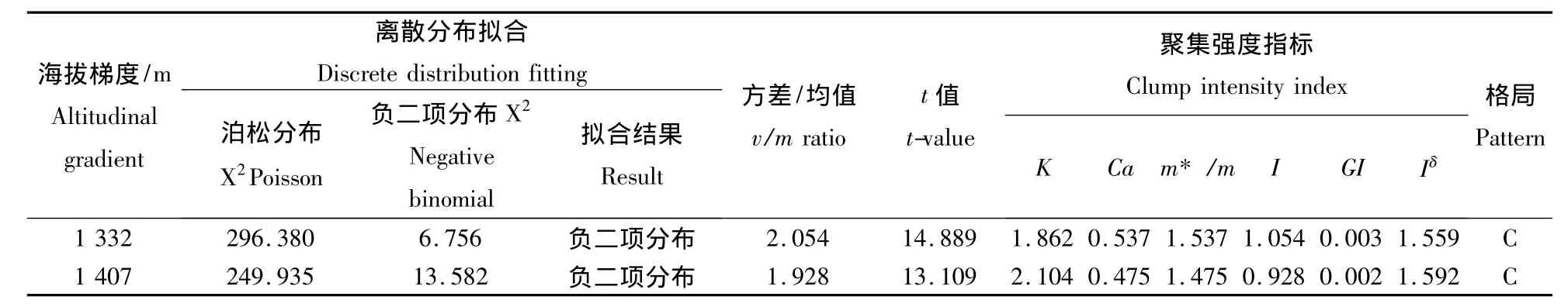
表3 不同海拔梯度宝天曼自然保护区锐齿栎种群的空间分布格局Table3 Spatial distribution pattern of Quercus aliena var.Acutesrrata population in Baotianman Nature Reserve at different altitudes
4 结论与讨论
1)本研究考察了宝天曼自然保护区锐齿栎种群在不同取样尺度上空间分布格局的变化情况,结果表明,在5种取样尺度上,锐齿栎种群均符合聚集分布类型,不同取样尺度上,种群反应的聚集程度不同,扩散型指数(Iδ)和聚块性指数(m*/m)随取样尺度的增大而减小,表明样方大小对宝天曼自然保护区锐齿栎种群空间分布格局的测定结果具有显著影响.这与宋萍等[9]研究天然黄山松(Pinus taiwanensis)的结果是一致的.在小尺度上,种群聚集度不高,而在大尺度上,聚集度明显增强,这与群落中幼树较少,中径树和大树较多有关,说明该地区锐齿栎种群正处于集群分布阶段,而且锐齿栎在宝天曼保护区为稳定种群,更新较好,并未有明显的退化现象.同时,本研究结果符合绝大多数自然种群都服从聚集分布而不服从随机分布的规律[12].
2)在宝天曼自然保护区不同发育阶段的锐齿栎种群空间分布格局在不同年龄阶段呈现不同的分布类型,幼树和中径树聚集程度较高,大树呈现随机分布.该地区锐齿栎种群的这种空间分布格局主要是由于种子的繁殖特性和生境条件的共同作用导致的.锐齿栎的种子为小坚果,成熟后主要靠重力落地,即使借助风力或山坡滚落也只能散布在母树周围不大的区域,幼苗呈聚集的蔟状分布.在种子和根茎的扩散过程中遇到障碍使大量繁殖体集中在一处而其它地方很少或完全没有;动物及人为活动对聚集分布也有加强作用.此外,锐齿栎种群密度大,能更好地改变微气候和小生境环境,提高了种群对不良环境的抵御能力,从而有利于种群的生长和存活.随着年龄的增加,作为一个喜光物种,锐齿栎幼树对光的需求逐渐增加,在同层次内,必须进入上层才能活的足够的光照.种内种间对光、水分和无机营养的竞争日趋激烈.很多幼苗和幼树被淘汰,存活下来的个体则呈现扩散趋势[3,17].对于中径树种群,种内和种间竞争进一步加强,随着个体的死亡,种群的聚集程度进一步下降,这有利于个体获得足够的资源.进入老龄期,不仅有竞争,而且由于生理衰老,导致个体死亡,种群聚集度再度下降[18],最终呈现随机分布类型.锐齿栎种群的空间分布格局在不同发育阶段的表现与种子传播、生境的异质性、种内和种间的竞争有密切的关系.不同发育阶段锐齿栎种群分布格局变化趋势表明对不同地区、不同年龄阶段的锐齿栎林应采取不同经营策略和管理措施.种群聚集度随年龄增长,在经营管理时应适时进行合理的人工疏伐,促进优势木的生长,从而获得最大森林生产效益.
3)海拔高度在1 332 m和1 407 m地区,锐齿栎种群空间分布格局均表现为聚集分布,但聚集程度不同,这主要是由于不同的海拔代表了不同的生境条件,地带性植被不同,水热条件及群落类型都存在差异.空间分布格局是植物种群研究的重要内容,是种群的重要结构特征之一,也是了解种群特征、种间关系以及与环境关系的重要手段[19].种群空间分布格局是生物群落中各种内外因素相互作用最直接的反应结果,不仅与物种的生物学特性和种间的竞争排斥有关,而且与物种的生境也有密切的联系[19~21].宝天曼自然保护区不同取样尺度、不同发育阶段、不同海拔高度的锐齿栎种群空间分布格局存在差异的成因与环境因子有着密切的关系,关于锐齿栎种群空间分布格局与环境因子的关系还有待于进一步的研究.
[1] 樊后保,王义弘.锐齿栎种群空间分布格局及动态的研究[J].福建林学院学报,1994,14(2):100-103.
[2] 李 博.普通生态学[M].呼和浩特:内蒙古大学出版社,1993:74-77.
[3] 张文辉,卢志军,李景侠,等.陕西不同林区栓皮栎种群空间分布格局及动态的比较研究[J].西北植物学报,2002,2(3):476-483.
[4] 张金屯.数量生态学[M].北京:科学出版社,2004.
[5] 马克明,祖元刚.兴安格叶松种群格局的分形特征—信息维数[J].生态学报,2000,20(2):188-192.
[6] 张志铭,朱学灵,刘晓静,等.宝天曼自然保护区栎林群落细根生物量研究[J].河南农业大学学报,2010,44(2):210-215.
[7] 兰国玉,雷瑞德.植物种群空间分布格局研究方法概述[J].西北林学院学报,2003,18(2):17-2l.
[8] 汤孟平,周国模,施拥军,等.天目山常绿阔叶林优势种群及其空间分布格局[J].植物生态学报,2006,30(5):743-752.
[9] 宋 萍,洪 伟,吴承祯,等.天然黄山松种群空问分布格局研究[J].江西农业大学学报,2008,30(5):830-832.
[10]宋 萍,洪 伟,吴承祯,等.珍稀濒危植物桫椤种群结构与动态研究[J].应用生态学报,2005,16(3):413-418.
[11]王本洋,余世孝.种群分布格局的多尺度分析[J].植物生态学报,29(2):235-241.
[12]郑元润.不同方法在沙地云杉种群分布格局分析中的适用性研究[J].林业科学,2001,37(1):6-10.
[13]蔡 飞,宋永昌.武夷山木荷种群结构和动态的研究[J].植物生态学报,1997,21(2):138-148.
[14]戚继忠,张 生.榛子种群空间格局及其动态的研究[J].吉林林学院学报,1996,12(2):99-103.
[15]王得祥,陈海滨.秦岭华山松种群分布格局及动态研究[J].西北植物学报,1998,18(4):622-628.
[16]刘仁林,朱玲琳.南方红豆杉天然群落种群空间分布规律的研究[J].环境与开发,1997,12(1):3-4.
[17]侯向阳,韩进轩.长白山红松林主要树种空间格局的模拟分析[J].植物生态学报,1997,21(3):242-249.
[18]韩 路,王海珍,周正立,等.塔里木胡杨林主要空间分布格局与动态研究[J].西南林学院学报,2007,27(2):1-4.
[19]张文辉,许晓波,周建云,等.濒危植物秦岭冷杉种群空间分布格局及动态[J].西北植物学报,2005,25(9):1840-1847.
[20] HOU J H,MI X C,LIU C R,et al.Spatial patterns and associations in a Quercusbetula forest in northern China[J].Journal of Vegetation Science,2004,15:407-414.
[21]罗天祥,李文华,赵士洞.中国油松林生产力格局与模拟[J].应用生态学报,1999,10(3):257-261.
