6种糖基化合物对台湾乳白蚁糖基水解酶分泌的影响
2011-04-04曾文慧刘瑞娴李志强刘炳荣李秋剑陈来泉钟俊鸿
曾文慧,刘瑞娴,李志强,刘炳荣,李秋剑,陈来泉,钟俊鸿*
(1.广东省昆虫研究所,广州 510260;2.广东韶能集团股份有限公司,韶关 510630)
木质纤维素 (lignocellulose)是植物细胞壁的主要组成成分 (Ohkuma,2003),是一类由纤维素(cellulose)、半纤维素 (hemicellulose)等多聚糖以及无定形态聚合物木质素 (lignin)结合在一起形成的混合物 (Arakawa et al.,2009),是地球上含量最丰富的可再生资源。
纤维素和半纤维素酶均属于糖基水解酶 (glycosyl hydrolase)蛋白家族。低等白蚁的木质纤维素水解酶系统包含三种不同类型的纤维素酶:外切葡聚糖酶,包括纤维二糖水解酶 (cellobilohydrolase,1,4-β -D -glucan cellobiohydrolase,CBH)和外葡萄糖水解酶 (exoglucohydrolase,1,4-β-D-glucan exoglucohydrolase)2种成份,内切葡聚糖酶 (endo-β-1,4-glucanases,EG,EC3.2.1.4)以及 β-葡萄糖苷酶 (β-glucosidase,BG,EC3.2.1.21)(Li et al.,2010)。同时,内切木聚糖酶 (endo-β-1,4-D-xylan,xylanohydrolase,EX,EC3.2.1.8)是低等白蚁重要的一类半纤维素水解酶 (Li and Sun,2001)。
研究发现低等白蚁拥有两个纤维素水解酶系统:一个在前肠 (Foregut)/唾液腺 (Salivary gland) +中肠 (Midgut),为内源性水解酶系统;另一个在后肠 (Hindgut),为后肠共生微生物水解酶系统 (Nakashima et al.,2002;Zhou et al.,2007)。但是,至今为止,低等白蚁的两个纤维素酶水解系统的调控机制还未有相关的研究报道。相较而言,在丝状真菌中已经发现了木聚糖酶和纤维素酶启动子结合型转录调控因子Xyrl、ACEI及转录激活因子X1nR等 (Ling et al.,2009;Stricker et al.,2008;Saloheimo et al.,2000)。研究发现真菌木质纤维素酶可以通过添加槐糖 (sophorose)、纤维二糖 (cellobiose)及乳糖 (lactose)等寡聚糖至培养基中作为调节因子来调节其分泌量 (Morikawa et al.,1995)。
本实验通过借鉴真菌纤维素酶和半纤维素酶分泌的调节因子,利用各种糖基化合物活体诱导台湾乳白蚁Coptotermes formosanus,研究它们对台湾乳白蚁4种糖基水解酶分泌的影响。通过测定EG、BG和CBH 3种纤维素酶及EX 1种半纤维素酶活力,来检测诱导效果。这些研究将给白蚁木质纤维素酶系的调控机制,提供重要的实验数据和理论依据。
1 材料与方法
1.1 实验材料
1.1.1 实验白蚁
供试虫源采自广州龙洞的白蚁诱捕箱,带回实验室饲养,挑取活力较好的台湾乳白蚁工蚁进行试验。
1.1.2 实验试剂
本实验选用的诱导试剂:木质素 (Lignin,low sulfonate content,Sigma),羧甲基纤维素钠(CMC-Na,天津福晨化学试剂厂),L-山梨糖(L-sorbose,Genview),D-果糖 (D-Fructose,Genview),葡萄糖 (Glucose,广州化学试剂厂),D-半乳糖 (D-Galactose,Genview);其它化学试剂:滤纸 (双圈定性滤纸),D-水杨苷 (DSalicin,上海晶纯试剂有限公司),对硝基苯-8-D-纤维二糖苷 (p-NPC,Sigma),木聚糖 (Xylan from Birchwood,Sigma),D-木糖 (D-Xylose,Genview),对硝基苯酚 (p-NP,Genview),牛清血蛋白组分 V(BSA,Albumin,Fraction V,Genview),3,5-二硝基水杨酸 (DNS,Genview),考马斯亮蓝 G-250(Coomassie brilliant blue G -250,Genview)。
1.2 实验方法
1.2.1 乳白蚁诱导培养
采用蛭石作为保湿材料,用灭菌蒸馏水湿润后均匀铺到直径为10 cm的培养皿中,每个培养皿放入150头供试乳白蚁工蚁,加盖培养。将6种诱导试剂分别配成1% (W/V)浓度溶液,各取约300 μL湿润滤纸,再放入加有白蚁的培养皿中,共六组。将装有白蚁的培养皿放入28℃恒温培养箱 (南京实验仪器厂,WMK-02型),每天添加一次诱导试剂保持湿润状态。在乳白蚁接触到含有各种诱导试剂的滤纸开始后的4 h、8 h、24 h、2 d、3 d、4 d、6 d和9 d取样。将乳白蚁肠道解剖成乳白蚁的前肠/唾液腺+中肠及后肠两部份。前肠/唾液腺+中肠部分用于检测白蚁内源性糖基水解酶活性变化,后肠部分用于检测白蚁共生微生物来源糖基水解酶活性变化,整头白蚁酶活力测定用作指示各种酶活性大小之间的比例变化。以接触诱导物之前的同批次乳白蚁作为参照,参照活性设定为100% 。
1.2.2 粗酶制备
每次取样选取各组中活力好的乳白蚁约15头置于0.9% (W/V)灭菌生理盐水中反复漂洗,之后用滤纸吸干表面水分,放入已灭菌的干净培养皿中。在实体解剖镜下将白蚁肠道解剖,分成前肠/唾液腺+中肠和后肠两部分,与5头未解剖的乳白蚁共分成3组分别放入装有500 μL HACNaAC缓冲液 (SAB,0.1 M,pH5.6)的离心管中再转移至玻璃组织匀浆器,冰浴研磨;研磨完毕将匀浆液吸至离心管后定容至 500μL,12000 rpm,4℃冷冻离心 (Sigma,3K15)15 min,取上清,再次12000 rpm,4℃冷冻离心5 min,取上清即为粗酶液于-20℃保存待用。
1.2.3 蛋白质浓度的测定
采用考马斯亮蓝法进行测定。将粗酶液按一定的比例进行稀释,取50 μL稀释后粗酶液,加入350 μL考马斯亮蓝G-250显色试剂,多功能酶标仪595 nm 比色 (PerkinElmer,1420-012 victor3),测定OD595,用蛋白质标准曲线求蛋白质含量。
1.2.4 内切-β-1,4-葡聚糖酶,β-葡萄糖苷酶及内切-β-1,4-木聚糖酶活力的测定
内切-β-1,4-葡聚糖酶 (EG),β-葡萄糖苷酶 (BG)以及内切 -β-1,4-木聚糖酶(EX)活力的测定,参照 Miller方法 (Miller,1959),均采用还原糖法测定。在1.5 mL离心管中,分别加入120 μL 1%CMC-Na,Salicin,Xylan,预热5 min,然后加入酶液 12 μL,37℃ 准确反应60 min,立刻加入120 μL DNS溶液终止反应,沸水浴 5 min,冰浴冷却,540 nm比色,测定OD540。EG和BG从葡萄糖标准曲线求得葡萄糖含量,计算酶活力单位;XE用木糖标准曲线求木糖含量,计算酶活力单位。EG,BG,EX酶活力U定义为,每毫克蛋白质在37℃,pH=5.6反应条件下分解底物,每分钟可以产生还原糖的量,表示为U/mg。每个酶切反应重复3次。
1.2.5 FPA滤纸酶活测定
采用还原糖法测定,用打孔器将滤纸制作成直径为5 mm的滤纸片,每个离心管中放入1份滤纸片,再加入120 μL SAB缓冲液 (pH 5.6)将其浸润。加入酶液 12 μL,37℃ 准确反应 60 min,然后加入120 μL DNS溶液终止反应,沸水浴5 min,冰浴冷却后,540 nm比色,测定OD540。葡萄糖标准曲线求得葡萄糖含量,酶活力单位计算同1.2.4。每个酶切反应重复3次。
1.2.6 纤维二糖水解酶 (CBH)活力测定
在1.5 mL离心管仲加入120 μL 1 mmol pNPC底物,预热5 min,然后加入酶液12 μL,37℃ 准确反应60 min,加入120 μL Na2CO3(0.6mol/L)溶液终止反应,405 nm比色,测定OD405,从p-NP标准曲线求得p-NP含量,计算酶活力单位。CBH酶活力U定义为:每毫克蛋白质在37℃,pH=5.6反应条件下分解pNPC,每分钟可以产生p-NP的量,表示为 U/mg。每个酶切反应重复3次。
2 结果与分析
2.1 不同糖基化合物对台湾乳白蚁内源性以及共生微生物来源的糖基水解酶分泌量的影响
选取用于对台湾乳白蚁进行活体诱导的化合物分为2类,第一类为大分子聚合物:木质素和羧甲基纤维素钠,第二类为单糖:山梨糖,果糖,葡萄糖。酶活力测定结果表明,这2类物质对乳白蚁后肠共生微生物来源的糖基水解酶的分泌与它们对内源性糖基水解酶的分泌作用相反。后肠部分:6种化合物对乳白蚁后肠EG,BG,CBH,EX 4种酶在整个实验时间内起到的基本为抑制分泌的作用,其中对后肠抑制作用最大的物质为果糖及山梨糖,只有 1% 半乳糖 (4 h,135%,0.540 U/mg)对滤纸分解有微弱的促进作用 (图1 f~j)。前肠/唾液腺+中肠部分:一方面山梨糖和果糖分别是为 EG(6day,126%,1.538 U/mg)与EX(6day,341%,5.146 U/mg)的最佳诱导物,同时山梨糖对BG(6day,147%,4.282 U/mg)和EX(6day,258%,3.892 U/mg),果糖对CBH(6day,249%,1.128 U/mg)也有较好的诱导效果。另一方面半乳糖和葡萄糖对内源性糖基水解酶促进作用不明显,且在大部分时间点对其分泌有抑制作用。实验结果还表明大分子物质木质素对4种内源性水解酶平均促进作用最大。它不仅为 FPA (6 day,149%,0.750 U/mg),BG(6day,178%,5.170 U/mg) 及 CBH(6 day,265%,1.201 U/mg)的最佳诱导物,其次对EG(6day,122%,1.486 U/mg) 和EX(6day,232%,3.505 U/mg)都具有相对好的诱导效果。五种酶活测定指标比较,CBH和EX的分泌对各种糖基化合物作用最为敏感且增幅最大。8个取样点内各种酶活力在4小时和第6天有高峰促进作用(图1 a~e)。

图1 台湾乳白蚁肠道不同部位糖基水解酶活性化学诱导曲线Fig.1 Graphs for chemical induced activity variations of glysosyl hydrolase in different parts of C.formosanus intestine
2.2 糖基化合物对台湾乳白蚁纤维素酶、半纤维素酶之间活性大小比例的影响
选取诱导后4 h(酶活高点)、3 d(酶活低点)、6 d(酶活高点)、9 d(酶活低点)的整头乳白蚁为样品,测定4种水解酶活性大小,以未接触诱导物的同批台湾乳白蚁作对照。对整头白蚁的酶活测定表明 (图2),EX的活力明显高于其它3种,是总酶活大小变化的主要影响因素。4种酶活力诱导最大值的大小顺序为 EX(6 d,9.356 U/mg) >BG(2 d,3.313 U/mg) > EG(2 d,1.541 U/mg) > CBH(6 d,1.476 U/mg)。分泌量有所提高的1% 木质素,1%CMC和1%山梨糖样品其整头白蚁4种酶活力的低点和高点的酶活大小比例均与对照白蚁基本一致,即EX:(BG+EG+CBH) 约为 2∶1。
3 结论与讨论
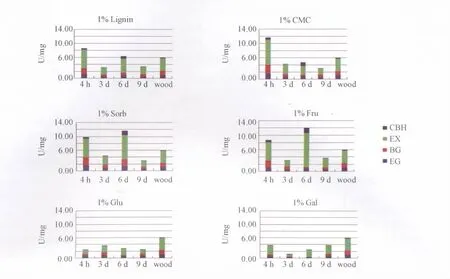
图2 台湾乳白蚁糖基水解酶肠道活性比例分布变化化学诱导曲线Fig.2 Graphs for chemical induced proportion variations of intestinal glysosyl hydrolase activities of C.formosanus
在T.reesei的纤维素调控机制中,T.reesei的生长营养条件能够显著影响其纤维素和半纤维素酶的产量,碳水化合物及他们的衍生物可以作为诱导物质促进其大部分纤维素酶的分泌 (Ling et al.,2009)。根据碳水化合物在细胞代谢中的一些共同中间产物,本研究选取用于对台湾乳白蚁进行活体诱导的化合物包括:代谢起始物质 (大分子聚合物)木质素和羧甲基纤维素钠;代谢下游产物 (单糖)山梨糖、果糖、葡萄糖和半乳糖。实验结果表明这些糖基化合物对乳白蚁不用来源的纤维素酶的分泌起着大相径庭的作用。
在6种化合物的影响下,乳白蚁后肠4种酶的分泌在8个取样点大部分为抑制状态,山梨糖和果糖的抑制作用最为明显,只有半乳糖对FPA活力有微弱的促进作用。而对于乳白蚁内源性糖基水解酶,木质素、山梨糖和果糖对酶的分泌起到了正向的诱导作用。
4种单糖中山梨糖和果糖互为差向异构体均为己酮糖,它们与己醛糖葡萄糖、半乳糖对后肠的作用与其对前肠/唾液腺+中肠的作用相反。丝状真菌中葡萄糖作为降解物,可以对纤维素酶分泌的造成反馈抑制,但是当葡萄糖耗尽之后纤维素则可以正常的表达 (Ilmen et al.,1997)。在本实验中葡萄糖对乳白蚁木质纤维素分泌的抑制作用,很可能也是由于代谢物的反馈抑制作用,半乳糖作为葡萄糖的差向异构体而产生了类似的作用。山梨糖在T.reesei中已经被证明可以通过启动子诱导糖基水解酶的分泌 (Ling et al.,2009),因此同理山梨糖的差像异构体果糖也很可能可以产生诱导作用。山梨糖和果糖对乳白蚁糖基水解酶的分泌表现出的诱导作用,可以则猜测为其可能拥有类似于T.reesei中控制纤维素酶和半纤维素酶的启动子,这些启动子的上下游序列中存在山梨糖等物质的激活位点。但是山梨糖和果糖对于后肠4种糖基水解酶的强烈抑制作用机理还需要进一步的研究。
木质素对乳白蚁内源性糖基水解酶5种酶活力指标的平均促进作用最大。木质素是一种高度分支的由多种芳香族羧酸组成的高分子物质,性质稳定降解缓慢 (Xie et al.,2000)。木质素通常在细菌或真菌的氧化酶作用下转变为芳环自由基和醌类才能进一步分解,而纤维素的降解产物可以为木质素的降解提供还原力 (Sewalt et al.,1996)。虽然木质素代谢与乳白蚁纤维素、半纤维素酶分泌之间的关系目前未见报道,但是Tartar等(2009)研究表明黄胸散白蚁Reticulitermes flavipes的酚氧化酶几乎全部来源于前肠和唾液腺,而且喂食木质素可以提高酚氧化酶的活性。结合本研究结果,木质素的作用机制可能为,由木质素作为一种间接促进酚氧化酶分泌的化学信号,刺激糖基水解酶分泌用来降解纤维素,从而产生各种代谢物质来直接诱导分解木质素的酚氧化酶的生产,即相当于提高了糖基水解酶的分泌量。
实验结果显示乳白蚁CBH和EX的分泌对各种糖基化合物作用最为敏感,其次为BG,以EG最不敏感,且后肠EG抑制最为强烈。所测定的乳白蚁4种糖基水解酶中,EX的活力明显高于其它三种,4种酶的大小顺序为EX >BG>EG>CBH。到目前为止,乳白蚁CBH的基因只在后肠的共生鞭毛虫中发现,而本实验中乳白蚁内源性CBH活性在接触诱导物后出现了大量的提高,因此推测乳白蚁是有可能存在自身基因组编码的CBH基因。木质素和半纤维素可以形成致密的网络结构,包围和加固纤维素与半纤维素骨架,将纤维素成份紧密的包埋起来 (Yu,2008),所以EX的大量分泌是木制纤维素代谢的基础。当EX分泌大量增加时,从木质纤维素中释放出大量纤维素,纤维素的结晶部份需要经过CBH的作用才能继续被EG和BG分解,这是其中一种可能的解释。因此EX的分泌量的增大伴随着CBH分泌量的相应提高,而BG和EG则由于处于代谢产物反馈调节中更加下游的位置使得其诱导不明显或出现抑制作用。同时在对T.reesei cbh1启动子的研究中发现,cbh1启动子上游区域存在葡萄糖抑制位点,可以在槐糖山梨糖醇的培养基中强烈诱导cbh1基因 (Takashima et al.,1996;Ilmen et al.,1996)。在本实验中葡萄糖和山梨糖对CBH也出现了相应的抑制和诱导作用,那么乳白蚁中是否存在类似与cbh1基因的调节机制还需要大量的基础研究工作。
此外,8个取样时间点内各种酶活力均没有呈现规律性的变化,只在4 h和6 d两个取样点有高峰促进作用。有研究表明,真菌纤维素酶的诱导合成与诱导物的浓度密切相关 (Sun,2007),因此诱导物对白蚁木质纤维素酶分泌的诱导作用的波动,可能反映了实际起作用的诱导物浓度的变化,相关的影响因子有待进一步研究。
References)
Arakawa G,Watanabe H,Yamasaki H,Maekawa H,Tokuda G,2009.Purification and molecular cloning of xylanases from the woodfeeding termite,Coptotermes formosanus Shiraki.Biosci.Biotechnol.Biochem.,73:710-718.
Ilmen M,Saloheimo A,Onnela ML,Penttila ME,1997.Regulation of cellulase gene expression in the filamentous fungus Trichoderma reesei.Appl.Environ.Microbiol.,63:1298-1306.
Ilmen M,Thrane C,Penttila M,1996.The glucose repressor gene cre1 of Trichoderma:isolation and expression of a full-length and a truncated mutant form.Mol.Gen.Genet.,251:451-460.
Li JF,SunY,2001.Studies on the characteristics of xylanase.Journal of Zhejiang University(Agric. & Life Sci.),27(1):103-106.[李卫芬,李建义,2001.浙江大学学报 (农业与生命科学版),27(1):103-106]
Li XH,Yang HJ,Ro B,Park EY,Jiang LJ,Wang D,Miao YG,2010.Enhanced cellulase production of the Trichoderma viride mutated by microwave and ultraviolet.Microbiol.Res.,165:190-198.
Ling M,Qin YLN,Liang Z,2009.Binding of two transcriptional factors,Xyr1 and ACEI,in the promoter region of cellulase cbh1 gene.Biotechnol.Lett.,227-231.
Miller GL,1959.Use of dinitrosalicylic acid reagent for determination of reducing sugar.Anal Chem.,3:426-428.
Morikawa Y,Ohashi T,Mantani O,1995.Cellulase induction by lactosein Trichoderma reesei PC-37.Appl.Microbiol.Biotechnol.,106-111.
Nakashima K,Watanabe H,Saitoh H,Tokuda G,Azuma JI,2002.Dual cellulose-digesting system of the wood-feeding termite,Coptotermes formosanus Shiraki.Insect Biochem.Mol.Biol.,32:777-784.
Ohkuma M,2003.Termite symbiotic systems:efficient bio-recycling of lignocellulose.Appl.Microbiol.Biotechnol.,61:1 -9.
Saloheimo A,Aro N,Ilmen M,Penttila M,2000.Isolation of the ace1 gene encoding a Cys(2)-His(2)transcription factor involved in regulation of activity of the cellulase promoter cbh1 of Trichoderma reesei.J.Biol.Chem.,275:5817-5825.
Sewalt VJH,Glasser WG,Fontenot JP,Allen VG,1996.Lignin impact on fibre degradation.1.Quinone methide intermediates formed from lignin during in vitro fermentation of corn stover.Journal of the Science of Food and Agriculture,71:195-203.
Stricker AR,Mach RL,de Graaff LH,2008.Regulation of transcription of cellulases and hemicellulases-encoding genes in Aspergillus niger and Hypocrea jecorina(Trichoderma reesei).Appl.Microbiol.Biotechnol,78:211 -220.
Sun XY,2007.Studies on the Synthesis Regulation of Lignocellulosesdegrading Enzymes in Penicillium decumbens.PhD Dissertation,Shandong Sniversity.[孙宪昀,2007.斜卧青霉木质纤维素酶系的合成调控研究.山东大学博士学位论文]
Takashima S,Iikura H,Nakamura A,Masaki H,Uozumi T,1996.A-nalysis of Cre1 binding sites in the Trichoderma reesei cbh1 upstream region.FEMS Microbiol.Lett.,145:361-366.
Tartar A,Wheeler MM,Zhou X,Coy MR,Boucias DG,Scharf ME,2009.Parallel metatranscriptome analyses of host and symbiont gene expression in the gut of the termite Reticulitermes flavipes.Biotechnol.Biofuels,2:25.
Tokuda G,Lo N,Watanabe H,Arakawa G,Matsumoto T,Noda H,2004.Major alteration of the expression site of endogenous cellulases in members of an apical termite lineage.Mol.Ecol.,13:3219-3228.
Xie Y,Yasuda S,Wu H,Liu H,2000.Analysis of the structure of lignin-carbohydrate complexes by the specific13C tracer method.Journal of Wood Science,46:130 -136.
Yu J,2008.Effects of Different Pretreatment Methods on Rice Hull for Enzymatic Hydrolysis.PhD Dissertation,[余君,2008.不同预处理工艺对稻壳纤维素酶酶解效果的影响.华中农业大学博士学位论文]
Zhou X,Smith JA,Oi FM,Koehler PG,Bennett GW,Scharf ME,2007.Correlation of cellulase gene expression and cellulolytic activity throughout the gut of the termite Reticulitermes flavipes.Gene,395:29-39.
