人胰腺细胞cDNA文库中丙型肝炎病毒NS4B结合蛋白基因的筛选
2011-03-09田梅梅张锦前
任 娜,吴 璐,李 贲,张 婷,田梅梅,张锦前,成 军,李 鑫
首都医科大学附属北京地坛医院,北京 100015
既往相关研究发现,HCV感染与糖尿病、移植后糖尿病、地中海贫血患者伴发的糖尿病或糖代谢异常均有相关性[1]。而且,肝脂肪变的存在与慢性丙型肝炎(CHC)的疾病进展有关[2]。Mason等[3]指出丙型肝炎病毒(HCV)感染与糖尿病(DM)密切相关。HCV包膜糖蛋白NS4B是一种多功能蛋白质,与HCV感染的慢性化和抗病毒免疫关系密切。因此我们应用酵母双杂交技术筛选人胰腺细胞cDNA文库中HCV NS4B的结合蛋白基因,为研究HCV慢性感染与胰岛素抵抗(insulin resistance,IR)、肝脏脂肪变性、代谢综合征(metabolic syndrome,MS)之间的相关性及其作用机制提供研究依据和方向。
1 材料与方法
1.1 材料 主要试剂及培养基:YPDA SD/-Trp、SD/-Leu、SD/-Trp/-Leu、SD/-Trp/-Leu/-His及SD/-Trp/-Leu/-His/-Ade培养基均购自美国Clonteeh公司。酵母双杂交系统3试剂盒:包括人胰腺细胞cDNA文库、pGADT7-AD克隆载体、pGBKT7-BD克隆载体、pGADT7-T对照质粒、pGBKT7-53对照质粒、酵母菌株AH109、酵母菌株Y187,购自美国Clonteeh公司。引物合成由北京奥科生物技术公司完成。
1.2 方法
1.2.1 酵母胰腺细胞cDNA文库的构建:按试剂盒说明书对文库进行扩增,纯化文库质粒后转化大肠埃希菌DH5α感受态细胞,随机挑取6个单克隆提取质粒, EcoRⅠ、XhoⅠ双酶切鉴定cDNA文库的多样性。然后将该文库转化酵母菌株 Y187,制备酵母胰腺细胞cDNA文库。提取新鲜生长的Y 187酵母菌落(直径>2mm)接种于150 mLYPDA液体培养基中,30℃振摇培养至A600>1.5时,把该培养物转摇至850 mL YPDA液体培养基中,30℃振摇培养至 A600=0.4~0.6。以醋酸锂法将胰腺细胞cDNA文库质粒(0.25 mg)转化入酵母菌株Y187进行文库转化,然后将转化物铺SD/-Leu板50块,30℃倒置温育7 d。同时将转化产物按 1∶10、1∶102、1∶104、1∶106和 1∶108分别铺于SD/-Leu培养基上检验转化效率。同时将pGADT7-AD和pGADT7-T对照质粒转化酵母菌株Y187。
1.2.2 诱饵质粒转化酵母菌株及鉴定:HCV NS4B蛋白的酵母表达载体pGBKT7-HCV NS4B由北京地坛医院传染病研究所构建,醋酸锂法转入酵母菌株AH109。转化后铺板于SD/-Trp固体培养基进行筛选,对直径>2mm的菌落随机挑取进行菌落PCR鉴定。同时将pGBKT7-BD和pGBKT7-53对照质粒转化酵母菌株AH109。
1.2.3 pGBKT7-HCV NS4B质粒转化的酵母菌株AH109与胰腺细胞cDNA文库质粒转化的酵母菌株Y187的配合:挑取新鲜生长的经菌落PCR鉴定正确的AH 109酵母菌落(直径>2 mm),接种于SD/-Trp液体培养基中,30℃振摇至A600为0.8~1.0。重悬于50m L 2×YPDA培养基中,与4m L含胰腺细胞cDNA文库的酵母细胞于摇床中30℃45 r/min(离心半径为5mm)振摇培养配合24 h。观察到三叶草状的二倍体细胞后,将其重悬于10 m L 0.25×YPDA培养基中,然后铺SD/-Trp/-Leu/-His平板和SD/-Trp/-Leu/-His/-Ade平板各25块,置30℃温箱培养。同时将配合产物按 1∶10、1∶102和 1∶103分别铺于 SD/-Trp/-Leu、SD/-Trp、SD/-Leu培养基上检验配合效率。生长16 d后把直径>2mm的菌落划线于铺有X-α-gal的SD/-Trp/-Leu/-His/-Ade培养基上,在此培养基上生长且变成蓝色的为阳性菌落。同时设立阳性对照和阴性对照,阳性对照为pGADT7-T质粒转化的酵母菌株Y187与pGBKT7-53质粒转化的酵母菌株AH109配合;阴性对照为pGBKT7-BD质粒转化的酵母菌株AH109与胰腺细胞文库质粒转化的酵母菌株 Y 187配合、pGBKT7-HCV NS4B质粒转化的酵母菌株AH 109、胰腺细胞cDNA文库质粒转化的酵母菌株Y187。
1.2.4 阳性质粒的分析:于SD/-Trp/-Leu/-His/-Ade固体培养基上挑取阳性菌落,于摇床中 30℃,250 r/ min(离心半径为5 mm)振摇5 d后用酸化玻璃珠法提取酵母质粒。电穿孔法将提取的质粒转化大肠埃希菌,并于含氨苄西林的 LB平板上培养,所获得菌落质粒用BglⅡ酶切鉴定后测序,测序结果与基因库中的序列进行比对分析。
2 结果
2.1 酵母胰腺细胞cDNA文库的成功构建及鉴定待转化的胰腺cDNA文库滴度在5×107CFU/mL左右,纯化后的质粒DNA浓度约为0.28 g/L。随机挑取文库菌落,提取质粒,用BglⅡ酶切鉴定胰腺细胞cDNA文库的多样性,结果显示插入片段大小不一,说明文库具有良好的多样性(见图1)。胰腺细胞cDNA文库转化酵母菌株 Y187后,检验转化效率,确定其滴度>1×106CFU/mL,确定胰腺细胞cDNA文库转化成功。
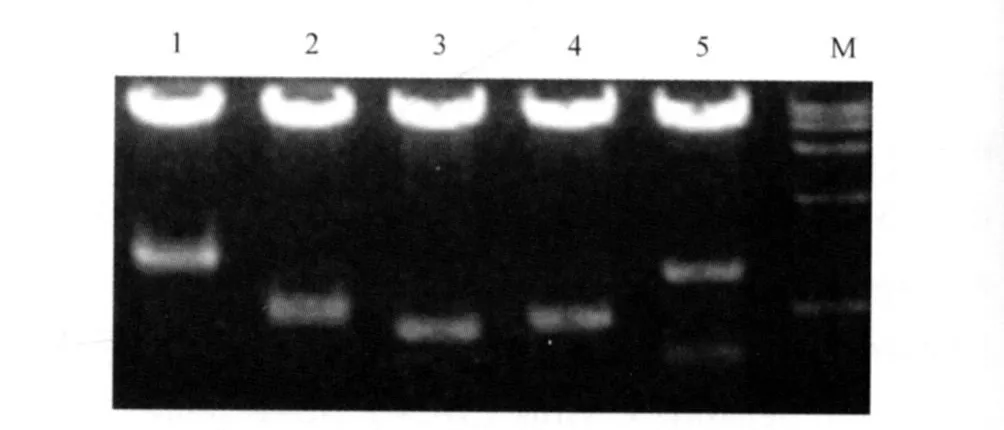
图1 部分文库质粒鉴定图 Lane 1~5为部分文库质粒BglⅡ酶切鉴定图;M:DNA MarkerFig 1 Several library o f identifying p lasm id
2.2 pGBKT7-HCV NS4B质粒转化AH109酵母菌株用醋酸锂法转化酵母细胞后在缺陷型SD-Trp培养基上筛选生长。培养4 d后,挑取菌落进行PCR扩增HCV NS4B基因,结果显示转化成功(见图2)。
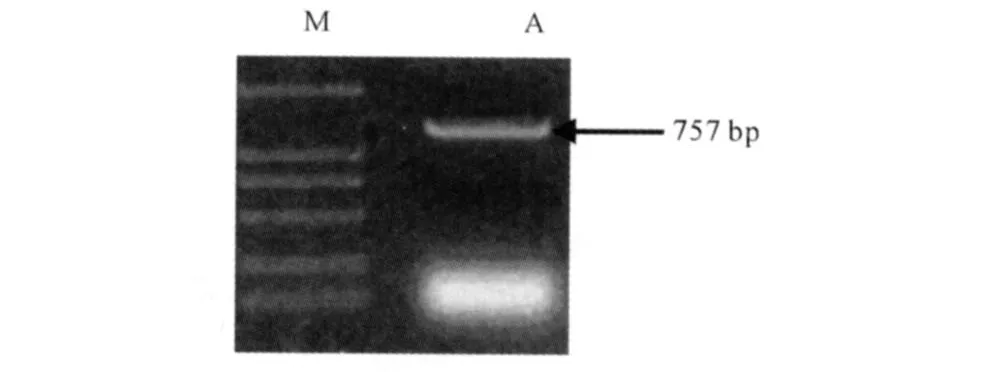
图2 pGBKT 7-HCV NS4B质粒转化AH 109酵母菌株后菌落PCR鉴定图 M:DNA Marker;A:pGBKT7-HCV NS4B质粒转化AH 109酵母菌株后单克隆菌株PCR扩增结果Fig 2 pGBKT7-HCV NS4B p lasm id transformation after AH 109 yeast strains co lony PCR appraisal figure
2.3 胰腺细胞cDNA 文库转化的酵母菌株Y187与pGBKT7-HCV NS4B质粒转化的AH109酵母菌株共同培养16 h后,用显微镜(40×)观察培养液可见配合生成的三叶草状二倍体酵母细胞(见图3)。
2.4 筛选出的胰腺细胞cDNA文库质粒测序与同源性分析初步结果 共筛选出 11个阳性克隆,其测序结果与基因库进行同源序列比对分析,发现HCV NS4B蛋白与 7种人类基因表达蛋白可能存在相互作用(见表1)。阴性对照无酵母菌落生长;阳性对照可见酵母菌落生长。

图 3 三叶草状二倍体酵母细胞图Fig 3 Clover-like diploid yeast cells
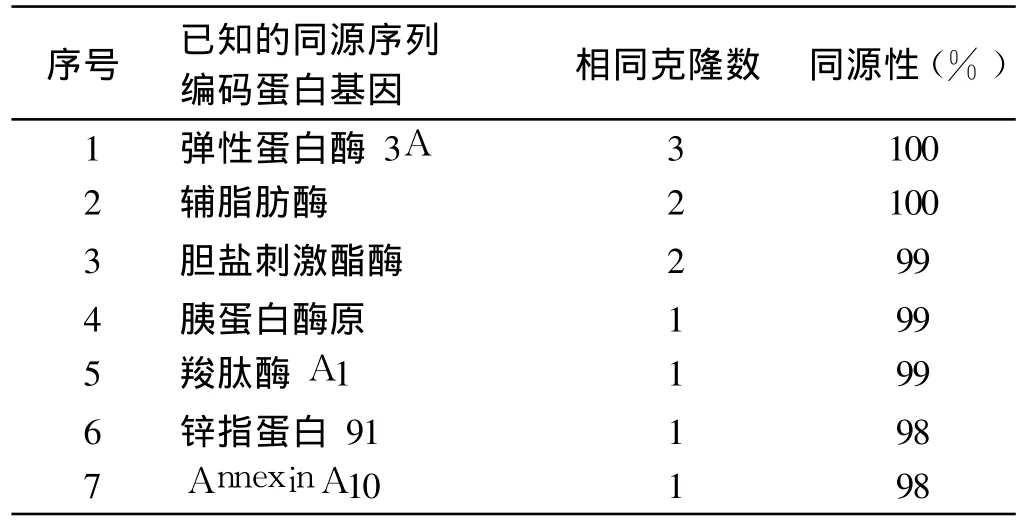
表1 阳性克隆测序后Blast比对结果Tab 1 Contrast resu lts after the positive cloning sequencing
3 讨论
国内外研究证实慢性丙型肝炎发病机制之一是肝脏脂肪变性,提示其可能是一种代谢性疾病,丙型肝炎病毒与MS密切相关,肝脏脂肪变性及IR可能是丙型肝炎病毒致代谢综合征的中心环节[4]。
HCV RNA与肝细胞脂类代谢物质的结合,可能是肝脏脂肪变形成的主要原因[6,7]。王琳等[8]运用酵母双杂交技术证实 HCV核心蛋白可与肝细胞中的载脂蛋白Apo-A1结合,它们的结合可能会影响Apo-A1的结构及其脂质转运的功能,进而影响脂类的代谢过程。通过对重组的E1/NS4B包膜蛋白与低密度脂蛋白(LDL)、极低密度脂蛋白(VLDL)和高密度脂蛋白(HDL)结合的分子特性进行研究,认为HCV进入血液后与脂滴结合形成复合物,通过LDL受体(LDL-R)进入肝脏,并进行复制,其产物与肝细胞中合成的载脂蛋白相互作用影响脂肪代谢,形成脂肪肝等[9,10]。
酵母双杂交系统是近年来新发展的分析蛋白-蛋白、蛋白-DNA、蛋白-RNA相互作用的一种有效的基因分析方法,为研究蛋白在体内生理情况下的相互作用提供了一种新的遗传学方法。我们选择以HCV NS4B作为酵母双杂交的诱饵蛋白,从人胰腺细胞cDNA文库中筛选与其相互作用的蛋白基因,寻找其中与糖、脂代谢相关的蛋白基因,为相关的机制研究奠定基础。
本实验将胰腺细胞cDNA文库质粒扩增后成功转化酵母菌株Y187,构建成功酵母胰腺细胞cDNA文库,以备进一步更广泛的酵母双杂交筛选工作使用。成功构建pGBKT7-HCV NS4B诱饵质粒,在酵母菌株AH109中表达HCV NS4B蛋白,并进一步与前述构建好的酵母胰腺细胞cDNA文库进行配合,筛选出可能与HCV NS4B蛋白存在相互作用的已知胰腺细胞蛋白基因 7种。这些蛋白以参与三大物质消化、吸收的酶类为主,其中参与糖、脂代谢的主要有弹性蛋白酶、辅脂肪酶和胆盐刺激酯酶。有研究表明,HCV可在胰腺细胞中复制,HCV NS4B在胰腺细胞中与前述这些蛋白结合后可能会影响它们的结构和功能,从而影响糖、脂代谢过程。
[1]Saliba F,Lakehal M,Pageaux GP,et al.Risk factors for new-onset diabetesmellitus following liver transplantation and impact of hepatitis C infection:an observational muhicenter study[J].Liver Transpl, 2007,13(1):136-144.
[2]SoresiM,TripiS,Franco V,et al.Impactof liver steatosison the antiviral response in thehepatitis C virusassociated chronic hepatitis[J]. Liver Intemat,2006,26(9):1119-1125.
[3]Mason AL,Lan JYN,Hoang N,et al.Association of diabetesmellitus and chronic hepatitis C virus infection[J].Hepatology,1999,29(2): 328-333.
[4]Zhang JQ,Fan XL.Relationship between chronic hepatitis C andmetabolic syndrome[J].World Chin J Digestol,2006,14(36): 3482-3486.
张锦前,范小玲.慢性丙型肝炎与代谢综合征[J].世界华人消化杂志,2006,14(36):3482-3486.
[5]Koike K.Hepatitis C as a metabolic disease:Implication for the pathogenesisof NASH[J].Hepatol Res,2005,33(2):145-150.
[6]Cortez-Pinto H.Concludingr emarks:metabolic syndrome,liver and HCV[J].Aliment Pharmacol Ther,2005,22 Supp1 2:83-85.
[7]Jan CF,Chen CJ,Chiu YH,et al.A population-based study investigating the association between metabolic syndrome and hepatitis B/C infection(Keelung Community-based Integrated Screening study No. 10)[J].Int JObes(Lond),2006,30(4):794-799.
[8]Wang L,Li K,Cheng J,et al.Interaction between hepatitis C virus core protein and apolipoprotein A 1[J].Word Chin JDigestol,2002, 10(9):1018-1021.
王琳,李克,成军,等.丙型肝炎病毒核心蛋白与载脂蛋白A 1结合的研究[J].世界华人消化杂志,2002,10(9):1018-1021.
[9]Sanyal AJ.Review artic le:non-alcoholic fatty liver disease and hepatitis C-risk factors c linical implications[J].Aliment Pharmacol Ther, 2005,22(2):48-51.
[10]Yoon EJ,Hu KQ.Hepatitis C virus(HCV)infection and hepatic steatosis[J].Int JMed Sci,2006,3(2):53-56.
