角膜混浊小鼠突变候选基因Map3k1克隆与序列分析*
2011-03-07吴刘成蒋荧梅王生存邵义祥
吴刘成,刘 春,蒋荧梅,王生存,邵义祥,*
(1.南通大学实验动物中心,江苏南通226001;2.南通大学比较医学研究所,江苏南通226001)
小鼠在正常发育过程中,胚胎期约在第11天开始出现原始的眼睑即单层细胞上皮,第12~17天,这层上皮细胞迅速发育分化形成复层上皮细胞,并出现角质化,在第14~16天之间眼睑不断生长发育覆盖了角膜,眼睑上缘和下缘融合,完成胚胎期眼睑闭合,出生后14 d眼睑重新开放[1]。眼睑开裂(Gaping lids gp)小鼠最早在1961年发现[2],由C57BL/6-ax品系的小鼠自发产生的常染色体隐性遗传突变模型,外显率100%,或者被称为lidgap-Gates(lg-Ga)模型,该突变系小鼠出生时眼睑开放,通过PCR技术发现lgGa突变系纯合子小鼠Map3k1基因第2~8外显子缺失,基因组缺失27.5 kb[3]。随着生物技术手段的发展,发现敲除Map3k1基因,角质形成性细胞微丝形成异常,细胞迁移力下降[4],研究表明Map3k1表达蛋白MEKK1通过激活JNK通路调控细胞的增殖、分化及迁移,基因敲除后JNK1磷酸化水平下降,细胞迁移障碍,胚胎时期眼睑不能够正常融合,出生时眼睑开放(eye open at birth,EOB)。
B6-Co(C57BL/6-Corneal opacity mouse,B6-Co)小鼠是本中心利用ENU诱变技术获得的具有角膜混浊表型的突变系小鼠,该突变系小鼠出生时眼睑开放,后角膜混浊[5]。蒋荧梅等利用SNP标记将B6-Co小鼠角膜混浊突变基因精确定位于13号染色体上112 546 283 bp~113 397 654 bp之间[6],Map3k1基因即位于定位区间,提出Map3k1作为B6-Co小鼠角膜混浊强力候选突变基因,本试验对该基因进行了克隆与序列分析。
1 材料与方法
1.1 材料
1.1.1 实验动物 C57BL/6(B6)小鼠、B6-Co突变系小鼠由南通大学实验动物中心提供,实验动物生产许可证:SCXK(苏)2008-0010。
1.1.2 主要仪器与试剂 冷冻离心机、PCR仪为德国Eppendorf公司产品;电泳仪为北京六一仪器厂产品;凝胶成像系统为江苏省捷达科技发展有限公司产品;RT试剂盒为Fermentas(MBI)公司产品;TaKaRa Ex Taq DNA Polymerase为宝生物工程(大连)有限公司产品;pGEM-T Easy Vector System试剂盒为Promega公司产品。
1.2 方法
1.2.1 组织RNA和DNA的提取 取小鼠大脑组织50 mg~100 mg,按照Trizol试剂盒说明提取总RNA;核酸定量仪测定RNA浓度;取2 μ L~5 μ L总RNA按照RT试剂盒说明逆转录合成cDNA。剪取小鼠尾尖1 cm,置于1.5 mL离心管;加入500 μ L消化液和蛋白酶K 6 μ L混合物,55℃水浴过夜;取过夜消化混合物,按照《分子克隆实验指南》操作[7]。
1.2.2 PCR体外扩增目的基因 在NCBI上查找小鼠Map3k1的mRNA序列,共20个外显子,全长6 963 bp,第1外显子481 bp,第20外显子2 581 bp,第2~19个外显子以cDNA产物为模板扩增,第1、20个外显子及上游调控序列以基因组为模板设计引物。将引物序列在小鼠基因组数据库中BLAST分析,确定引物的特异性。第2~20外显子共设计8对引物:Map3k1-2 F:GGAAGACGAGTGGT TGG,R:GGCTGTAGAAGTCATGGGAT,产物长度918 bp;Map3k1-3 F:CGGAACACCATCCAGAAGT T T,R:CAGCATGGCTCTCAATGT TT,产物长度777 bp;Map3k1-4 F:TCTATAGTCTGCGCTGACCCT,R:TCAGAAATGTCCTCCGAGCT,产物长度857 bp;Map3k1-5 F:AGAAAACAGCTCCCT TGAGCA,R:TAAGGCTGTT TCGCTT TGGT,产物长度1 302 bp;Map3k1-6 F:CGATGTCAGCGTCTCAGGAT,R:AGGCGAGATGAT TGGAGTGTT,产物长度798 bp;Map3k1-7 F:ACTGGGAGGCTATGAGGTTC,R:GT T TATCAGCCCAGTCTCGTAT,产物长度1 526 bp;Map3k1-8 F:TCTCTGTCACGACCCATCAC,R:CAGTGCTAATCTGACCGAATG,产物长度815 bp;Map3k1-9 F:GCACTCTTGCCAT TTACCG,R:T TCT TCT TGAGT T TCCTGCG,产物长度864 bp。Map3k1上游约5 kb序列共设计4对引物:Map3k1-01 F:CGCCAGGTAGTGCGAGAC,R:GGCAGCTTTATCCCGT TG,产物长度759 bp;Map3k1-02 F:CTCCCTGGTTGCCCTGT,R:TCCT TAGCCAATCTGGTCCT,产物长度1 193 bp;Map3k1-03 F:TCTGCTGTCAT TATGCACCTAT,R:T TGTT TCT TTGCATGTAGGACT T,产物长度1 250 bp;Map3k1-04F:CACTT TCAGGCAGGGTCAG,R:GGCTGAACGCT TGGAGTC,产物长度1 038 bp。由于Map3k1第1外显子GC含量高达80.2%,按常规设计引物原则不能成功扩增,必须提高引物的退火温度:Map3k1-1.1 F:GCCCCGCTCGCT TCAT TCA,R:CCGCCCTGCCCATCTACTTCC,产物长度252 bp;Map3k1-1.2 F:CGGCGACGGGGAAGTAGATG,R:CGCAGGCACGAGCGAATGT,产物长度328 bp;这两对引物Tm值均为63℃。
PCR反应体系为10×PCR buffer(含Mg2+)5.0 μ L,dNTP(2.5 mmol/L)4.0 μ L,上游引物(10 μ mol/L)2 μ L,下游引物(10 μ mol/L)2 μ L,模板DNA,2.5 ng,Ex Taq DNA poly mease(5 U/μ L)0.25 μ L,补灭菌去离子水至50 μ L。扩增条件:根据引物Tm值和扩增长度设置退火温度、变性时间及延伸时间。第一外显子GC含量高,需要在普通Taq酶反应体系中添加甜菜碱才能成功扩增,终浓度至2.5 mol/L。反应条件:94℃5 min预变性质94℃1 min,63℃30 s,72℃30 s,共35循环,72℃5 min。根据引物1.1和1.2成功扩增出目的片段。取PCR终点产物,凝胶电泳及成像保存。
1.2.3 PCR产物的克隆 按照上海生工生物工程技术服务有限公司UNIQ-10柱式胶回收试剂盒操作纯化PCR产物;与T载体连接,4℃过夜;取5 μ L链接产物和50 μ L~100 μ L感受态细胞加入预冷的离心管中;冰浴5 min;42℃热休克90 s;冰浴30 min;加入200 μ L LB培养液,37℃、250 r/min振荡培养45 min;取120 μ L~200 μ L菌液涂于15 g/L琼脂LB平板(添加氨苄青霉素、X-Gal和IPTG);37℃培养过夜;挑取白斑,扩培摇菌12 h~14 h;质粒DNA分子提取;电泳检测;EcoRⅠ酶切释放目的片段,电泳检测,送上海生工生物工程技术服务有限公司测序。
1.2.4 序列分析 目的片段测序后,在小鼠基因库中BLAST测序的序列是否与正常组小鼠基因一致。如存在碱基的突变,查看其是否位于蛋白编码区,并预测在蛋白水平上氨基酸序列可能引起的变化。
2 结果
2.1 RT-PCR体外扩增目的基因
为了测序的需要,PCR扩增的DNA片段都在1 500 bp以下,扩增第2到第19个外显子时,以B6小鼠和B6-Co小鼠的cDNA为模板,在扩增第1和第20个外显子时,以B6小鼠和B6-Co小鼠的基因组DNA为模板,PCR产物经10 g/L的琼脂糖凝胶电泳观察结果,成功扩增出了Map3k1基因的8个片段以及上游约5 kb序列(图1和图2),第1外显子分段PCR结果见图3。
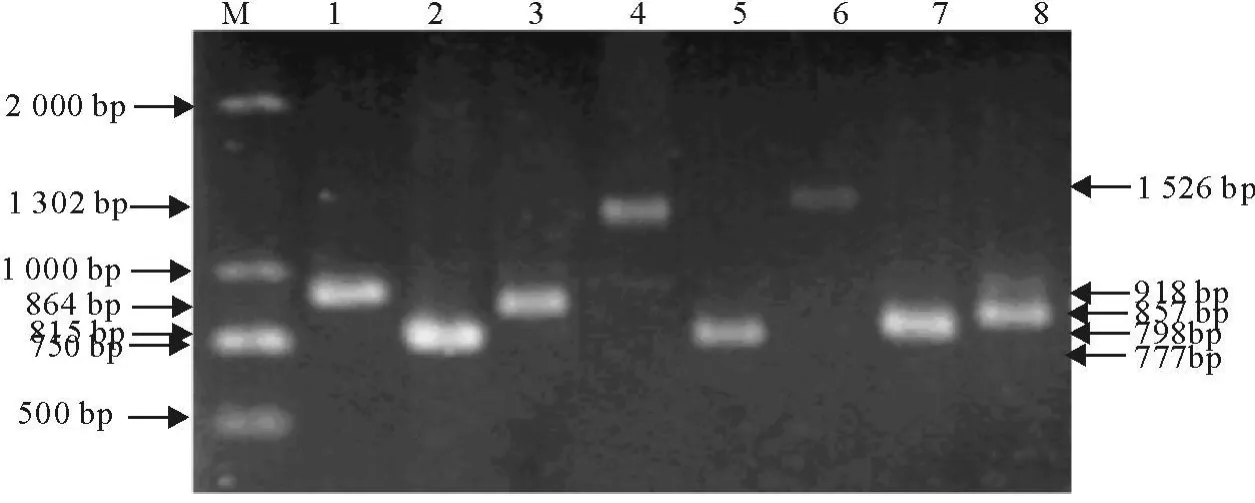
图1 Map3k1 RT-PCR产物电泳结果Fig.1 Map3k1 RT-PCR products electrophoresis result
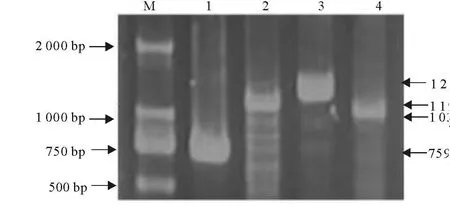
图2 Map3k1上游调控序列四段PCR产物电泳Fig.2 Four fragments of Map3k1 forward regulate PCR products electrophoresis

图3 Map3k1第1外显子分段PCR产物电泳 Fig.3 Fragments of Map3k1 first exon PCR product electrophoresis
2.2 质粒DNA的抽提、鉴定及酶切分析
将挑出的白斑扩增培养过夜,进行质粒提取,电泳检测(图4),每个泳道有3条带,泳动速度最快的是超螺旋状态的质粒,中间的是螺旋状态的质粒,最慢的是线型质粒DNA。
EcoRⅠ酶切反应结束后,凝胶电泳检测,由于T载体上插入片段两端有EcoRⅠ酶切位点,所以,酶切后质粒电泳一般有两条带,泳动速度最快的是目的片段,最慢的是被酶切的线性化的T载体(图5)。
2.3 测序结果分析
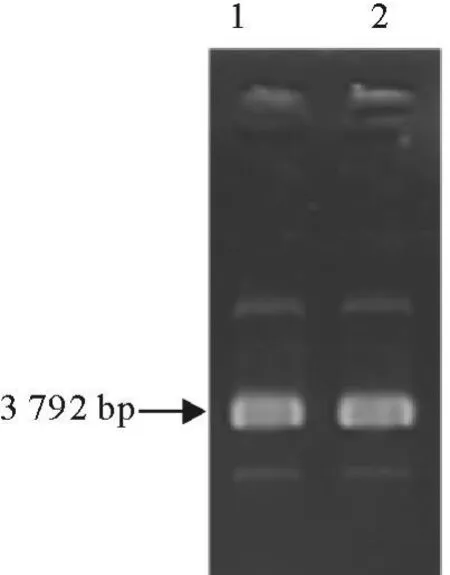
图4 部分重组质粒电泳结果Fig.4 Part of recombinant plasmid electrophoresis

图5 重组质粒酶切产物电泳Fig.5 Recombinant plasmid restriction enzyme digestion product electrophoresis
测序峰图显示,B6和B6-Co小鼠在第2外显子PCR产物测序峰图中均出现重叠峰,其他外显子及上游调控序列PCR产物测序峰图均没有出现重叠峰,克隆测序进一步验证该位点存在突变,与NCBI数据库比对,发现B6小鼠和B6-Co小鼠Map3k1基因mRNA序列第955位,即小鼠13号染色体112 559 574碱基与数据库比对发现,碱基T突变为A,这样编码的第314氨基酸由亮氨酸变为谷氨酸,蛋白质二级结构和跨膜区域均发生了变化。也有可能是数据库存在错误,还需要从多个实验动物种子中心引种,对该碱基位点进行验证。
3 讨论
3.1 Map3k1的功能
Map3k1表达的蛋白MEKK1具有Ser/Thr激酶活性,可以磷酸化与其结合的蛋白,介导蛋白质之间的相互作用并影响其行为和功能[9]。MEKK1可以被上游许多调控因子激活,如FAK、RhoGEF、RhoA、Rac、以及actinin protein等,从而活化下游相关的信号通路,因此可以和许多信号转导途径相联系,可以作用于伤口的愈合以及癌细胞的侵袭等细胞迁移相关事件[10]。MEKK1在体内高度选择性地磷酸化MKK4,从而激活JNK,JNK/SAPK接受上游信号被激活后,可以进一步使核内转录因子c-jun氨基末端丝氨酸残基磷酸化,激活c-jun而增强其转录活性[11]。
MEKK1是活化JNK的上游重要调控因子,MEKK1激酶结构域敲除后JNK1磷酸化水平明显降低,从而使该信号通路阻断,细胞微丝骨架异常,细胞迁移受阻。MEKK1在眼睑上皮细胞前一边缘高水平表达,并且需要F肌动蛋白的形成。除了MEKK1还需要TGF-β/activin诱导肌动蛋白张力纤维的形成,从而体外培养的角质形成细胞才能迁移,总之,MEKK1调节上皮层细胞运动促使眼睑闭合[12]。
3.2 B6-Co突变系小鼠候选基因的探讨
Zhang L等[4]对Map3k1激酶结构域敲除系小鼠进行研究表明,只有纯合的敲除系小鼠才表现为出生时眼睑开裂,杂合突变系表现基本正常,为隐性致病型,杂合子存在正常的Map3k1等位基因,可以满足Map3k1调控作用。B6-Co突变系小鼠出生时眼睑开裂,有单眼,也有双眼,发生率约为43.1%,为单基因显性遗传,但至目前也未获得纯合子,也未见胚胎致死。与单基因显性遗传规律还存在差异,其遗传机制尚需深入探讨。B6-Co小鼠眼睑发育缺陷导致角膜发育异常和先天性眼表疾病,该小鼠角膜混浊的病理过程呈典型的炎症反应,与人类角膜炎的病理变化过程相似[13],具有很高的开发价值。
通过克隆测序的方法能够比较准确地获得目的基因的DNA序列,有助于正确判断目的基因相关的分子机制[14-15]。本试验结果表明Map3k1基因存在单碱基突变,编码的氨基酸也发生了改变,这为进一步深入研究该基因对B6-Co小鼠眼睑发育及角膜混浊形成机制奠定基础。由于基因结构及调控机制的复杂性,非编码区对基因的功能也有重要作用,kitl1-bao纯合子小鼠kitl基因非编码区第8号内含子第2个碱基由T转换为C,从而引起mRNA剪接错误,导致基因功能发生了重大变化[16],所以要继续关注该基因的内含子、基因下游以及上游更远端的序列。Map3k1基因也有可能产生表达量的变化,或表达的蛋白质剪切加工过程中受到影响,因此,应进一步采用荧光定量PCR技术检测Map3k1基因的表达调控,用Western blot技术检测MEKK1蛋白水平的变化。
同时,在13号染色体60cM附近可能存在其他突变候选基因,从而导致了小鼠出生时眼睑开放的发生,也需要继续关注小鼠13号染色体60cM附近其他基因的功能。也有研究表明位于13号染色体75cM附近的Fgf10(fibroblast growth factor 10)基因突变后也能够产生相似的表型[17],虽然不在定位区间内,但遗传距离只是相对的,值得予以关注。
[1] Jin C,Yin F,Lin M,et al.GPR48 regulates epithelial cell proliferation and migration by activating EGFR during eyelid development[J].Invest Ophthalmol Vis Sci,2008,49(10):4245-53.
[2] Russell L B.lg-lid gap[J].Mouse News Lett,1961,25:64.
[3] Juriloff D M,Harris M J,Mah D G.The open-eyelid mutation,lidgap-gates,is an eight-exon deletion in the mouse Map3k1 gene[J].Genomics,2005,85(1):139-142.
[4] Zhang L,Wang W,Hayashi Y,et al.A role for MEK kinase 1 in TGF-β/activin-induced epithelium movement and embryonic eyelid closure[J].EMBO,2003,22(17):4443-4454.
[5] 唐建忠,朱昌来,邵义祥,等.遗传缺陷性小鼠角膜炎模型的超微病理观察[J].临床与实验病理学杂志,2008,24(5):593-595.
[6] 蒋荧梅,刘 春,吴刘成,等.SNP标记对角膜混浊小鼠突变相关基因的精细定位[J].遗传,2010,32(5):486-491.
[7] Sambrook J,Russell DW.分子克隆实验指南[M].3版.北京:科学出版社,2002:479-483.
[9] Xia Y,Wang J,Xu S,et al.MEKK1 mediates the ubiquitination and degradation of c-Jun in response to osmotic stress[J].Mol Cell Biol,2007,27(2):510-517.
[10] Cuevas B D,Abell A N,Johnson G L.Role of mitogen-activated protein kinase kinase kinase sinsignal integration[J].Oneogene,2007,26(22):3159-3171.
[11] Das K C,Muniyappa H.c-Jun-NH2 terminal kinase(JNK)-mediates AP-1 activation by thioredoxin:phosphorylation of cJun,JunB,and Fra-1[J].Mol Cell Biochem,2010,337(1-2):53-63.
[12] Takatori A,Geh E,Chen L,et al.Differential transmission of M EKK1 morphogenetic signals by JNK1 and JNK2[J].Development,2008,135(1):23-32.
[13] 刘 春,袁志云,杨菲菲,等.C57BL/6小鼠葡萄球菌性角膜感染模型的建立及相关研究[J].眼科研究,2010,28(3):199-202.
[14] 付 薇,陈进喜,覃芳芸,等.猪链球菌9型cps9G基因的克隆与序列分析[J].动物医学进展,2009,30(1):1-4.
[15] 屈 雷,闫海龙,庞全海,等.佳米驴防御素Abd-1基因克隆及序列分析[J].动物医学进展,2009,30(8):34-38.
[16] 薛整风,陈 兵,茅慧华,等.kitl非编码区突变导致RNA剪切异常的小鼠[J].动物学报,2007,53(2):332-338.
[17] Puk O,Esposito I,¨Oker T,et al.A new Fgf10 mutation in the mouse leads to atrophy of the harderian gland and slit-eye phenotype in heterozygotes:a novel model for dry-eye disease[J].Invest Ophthalmol Vis Sci,2009,50(9):4311-4318.
