松江鲈Myostatin基因内含子和线粒体D-loop DNA序列分析
2011-02-08费晓燕卢祥云徐建荣顾志良
邵 芳,费晓燕,卢祥云,徐建荣,顾志良
(常熟理工学院 生物与食品工程学院,江苏 常熟 215500)
松江鲈(Trachidermus fasciatus)属鲉形目,杜父鱼科,松江鲈属.近年来由于资源枯竭,种群濒危,已被列为国家二级保护动物.目前对松江鲈的研究主要集中在生态、生理、胚胎发育与繁殖习性等方面[1].为有效保护和合理利用松江鲈资源,对其种群遗传结构和遗传多样性的研究就显得尤为重要.
肌肉生长抑制素(Myostatin,MSTN)作为骨骼肌生长发育的负调节因子[2],引起生物学家的广泛关注.弄清MSTN基因的结构和功能对阐明骨骼肌生长发育的调控机理具有重要意义.研究表明鱼MSTN基因由三个外显子和两个内含子组成[3-5],我们对松江鲈MSTN基因的全长cDNA和组织表达特异性作了研究[6],但目前对松江鲈MSTN基因的内含子序列尚未有报道.mtDNA是独立于细胞核基因组外的环状双链DNA分子,在mtDNA上存在一个独特的D-loop结构,是整个线粒体基因组序列和长度变异最大的区域,其进化速度最快.线粒体DNA的D-loop区可用于研究物种起源和进化以及种内种群的系统进化分析,探讨物种在进化过程中可能发生的变异情况.鱼类mtDNA是一个相对独立的复制单位,具有分子小、结构简单、重组率低、进化速度快、不同区域进化速度有差异等特点,是鱼类进化遗传学、分子生态学、遗传多样性及其保护生物学等研究的重要标记[7].
本研究对MSTN基因内含子1、2以及长江、丹东和葫芦岛的松江鲈线粒体D-loop区扩增和测序,检测不同地理群体松江鲈线粒体D-loop区的多态性,探索种群内和种群间的遗传多样性状况,为今后松江鲈的资源保护、开发和养殖提供相应的理论依据.
1 材料与方法
1.1 实验材料:
本实验所用的松江鲈来自长江(C)、丹东(D)、葫芦岛(H)三个地理群体.实验所用材料由苏州市长江特色水产繁殖工程中心提供.将鱼宰杀后取肌肉组织在-80℃冰箱中保存备用,用酚-氯仿法提取基因组DNA.
1.2 引物设计与合成
根据我们克隆的松江鲈Myostatin基因cDNA序列(Genbank登录号:GU198192),以及斑马鱼和鲤鱼Myostatin基因DNA序列(Genbank登录号:AY693972,GQ214770),分别设计引物Sjl intr1 F1/R1,Sjl intr2 F1/R1以扩增松江鲈Myostatin基因的内含子1和内含子2序列.根据GenBank数据库中已公布松江鲈线粒体D-loop部分序列(Genbank登录号:AB188172),设计引物SjldpF1/R1,扩增松江鲈D-Loop部分片段,以分析不同地理群体的关系.引物由上海生工生物工程有限公司合成.

表1 实验所用的引物序列
1.3 松江鲈Myostatin基因内含子的PCR扩增和克隆
PCR扩增在25μL反应体系中进行:0.2μL ExTaq酶(5U·μL-1,TaKaRa Tokyo,Japan),2.5μL10×PCR ExTaq Buffer(with Mg2+),2μL dNTP(2.5mmol·L-1each),Sjl intr1 F1和Sjl intr1 R1各1μL(10μM),基因组DNA(100ng/μL)1μL,加灭菌水至25μL.PCR扩增条件为95℃变性3min;然后在95℃ 30 Sec,60℃30Sec,72℃1.5min,进行30次循环;72℃延伸10min;4℃保存.将PCR扩增产物于1%琼脂糖凝胶电泳,切胶回收目的片段.将回收片段克隆到PCR2.1 Vector(Invitrogen,美国)中,挑取阳性菌落,于LB液体培养基(Amp+)中摇菌培养,提取质粒,经鉴定后将重组质粒送往上海桑尼生物科技公司进行测序.
1.4 不同地理群体线粒体D-loop序列扩增和序列测定
PCR扩增反应体系和反应条件同上述1.3.将PCR产物进行琼脂糖凝胶电泳检测,回收纯化目的片段,直接测序.
1.5 数据分析
将获得的序列在DNAMAN5.0软件上进行分析,并进行物种间同源性比较.利用MEGA4.1软件中的Kimura 2-parameter法计算净遗传距离矩阵,以距离矩阵邻接法(Neighbor-Joining,NJ)建系统发生树,通过自引导检验(bootstrap)获得系统分支的置信度(重复次数为1000).
2 结果与分析
2.1 松江鲈MSTN基因内含子的扩增
以松江鲈基因组DNA为模板,用引物Sjl intr1F1/R1和Sjl intr2F1/R1进行PCR扩增,PCR产物在1%琼脂糖凝胶中电泳检测(图1).电泳检测结果显示,分别得到了450bp和1000bpPCR扩增产物,与设计的目的片段长度相同.将这两个片段分别回收后克隆到PCR2.1载体中,经测序后获得了两个片段的核苷酸序列,与松江鲈的MSTN基因cDNA序列比对,确认该序列为松江鲈MSTN基因的内含子1和内含子2序列.
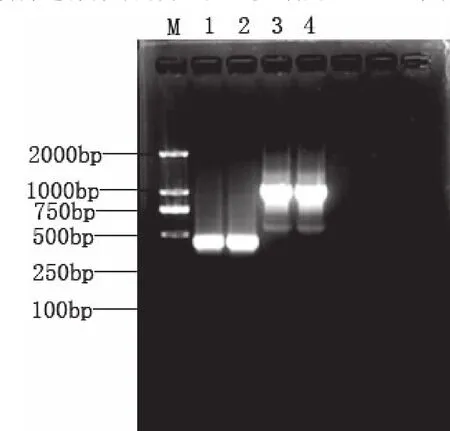
图1 松江鲈MSTN基因内含子PCR扩增产物的1%琼脂糖凝胶电泳图
2.2 松江鲈MSTN基因内含子的序列分析
通过PCR扩增并克隆测序后获得了松江鲈MSTN基因内含子1和内含子2的序列,内含子1长度为325bp,内含子2长度为835bp(图2和3),两个内含子的两端都符合GT-AG规则.在内含子2中发现了一个(AC)14的微卫星序列.

图2 松江鲈MSTN基因的内含子1序列

图3 松江鲈MSTN基因的内含子2序列
根据对其他物种MSTN基因序列的分析,发现鱼类中存在几种MSTN基因,包括MSTN1a,MSTN1b,MSTN2a,MSTN2b4种基因,且每个物种的MSTN基因都是含有两个内含子,三个外显子,不同物种的内含子长度不尽相同[6,8-10].表2为不同物种MSTN基因的内含子长度,可以发现有的物种中内含子1比内含子2长,有的物种内含子2比内含子1长.斑点叉尾鮰的MSTN内含子1最长为1748bp,瘤棘鲆MSTN内含子1最短为244bp,斑马鱼MSTN-2的内含子2最长为2201bp,尖吻鲈的MSTN内含子2最短为188bp.而我们得到的松江鲈MSTN基因的内含子2比内含子1长.
2.3 不同地理群体松江鲈的线粒体D-loop序列比较
对分别来自长江、丹东、葫芦岛三个不同地理群体的松江鲈18个个体线粒体D-loop片段进行了PCR扩增和测序.用DNAMAN软件对这18个个体进行序列比对后,共发现19个多态位点,但是未发现具有地理群体特征的单倍型.将这18个个体用MEGA软件绘制N-J进化树,也未发现三个地理群体松江鲈的特征性差异.
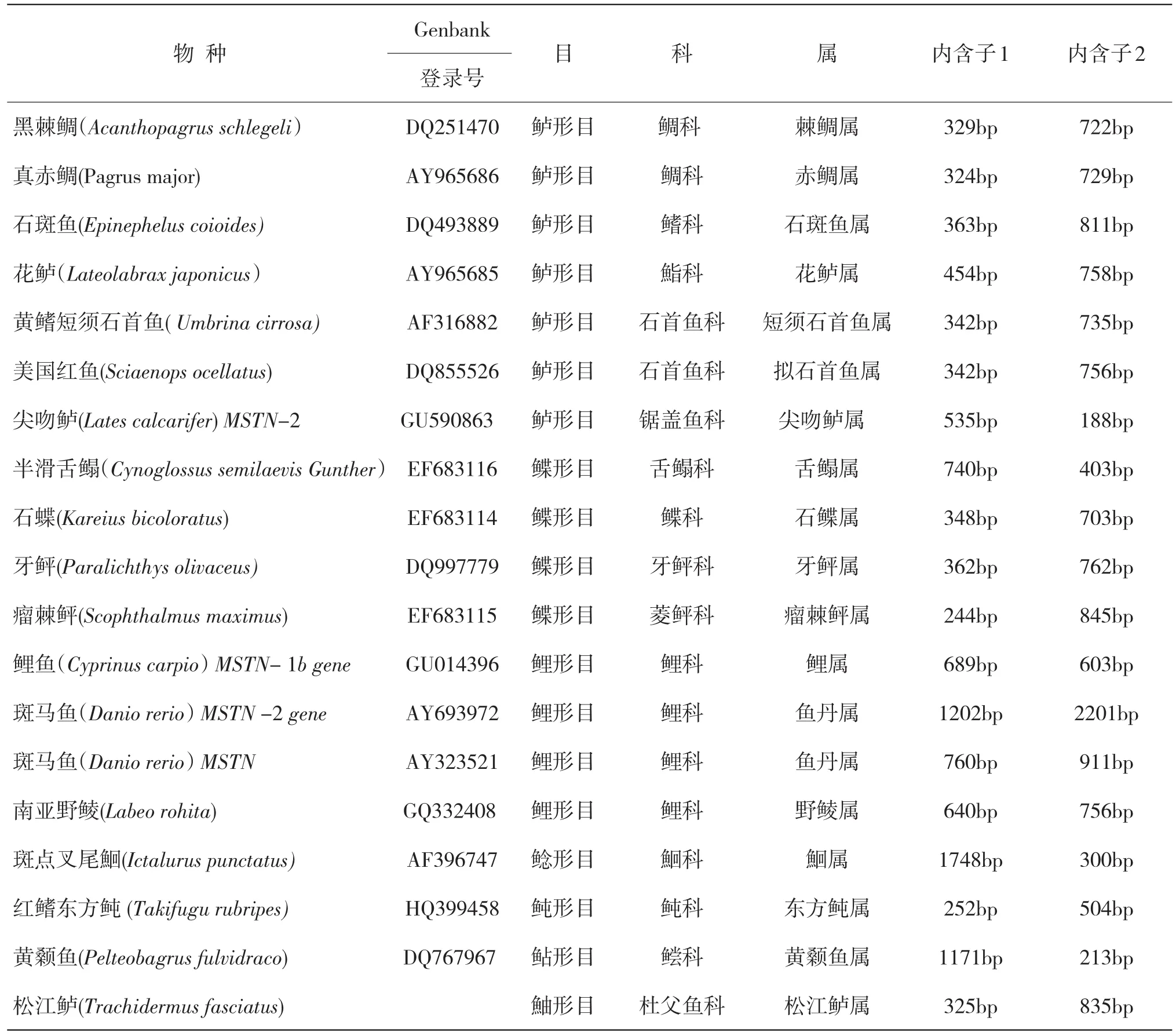
表2 不同物种MSTN基因内含子的长度比较
3 讨论
本研究根据松江鲈MSTN的cDNA序列设计引物,获得了松江鲈MSTN基因的内含子1和内含子2序列,该序列符合真核基因内含子的特征.在内含子2中发现了1个(AC)14的重复序列.研究表明,已在多种哺乳动物的MSTN基因中发现微卫星序列,De la Rosa-Reyna(2006)在牛MSTN内含子1中的T-motif中发现了(TG)微卫星[11];姜运良(2004)在猪MSTN侧翼区发现(TG)13的微卫星标记[12].在鱼类的MSTN基因中也发现了微卫星序列:金鲷MSTN内含子Ⅱ和3′-UTR区内都存在(AC)n重复序列[4],石鼓鱼MSTN序列的3′-UTR区存在(AC)29微卫星序列[13];大黄鱼肌肉生长抑制素基因的3′-UTR区存在(CA)n微卫星序列,且该微卫星座位与大黄鱼体长和体重存在轻度的正相关[14].
对物种进化的分子生物学研究目前主要集中在线粒体DNA方面,多数主要依赖于线粒体D-loop的信息,我们检测到了松江鲈不同个体间的D-Loop区序列存在差别,但是三个地理群体未发现明显的地理序列特征.可能是由于这些地理群体分隔时间短,没有明显大的差别.这一结果在对松江鲈MSTN基因序列多态性的检测时一定程度上得到了验证,在松江鲈MSTN基因的两个内含子和3′-UTR区未发现多态性位点(未发表资料),三个地理群体之间没有明显差异,一方面说明了MSTN基因比较保守,另一方面,可能也是由于这几个地理群体的松江鲈分隔时间短.我们将进一步检测其他线粒体基因中的变异情况,加入其他基因的信息,分析这些地理群体的松江鲈之间进化的关系.
[1]王金秋,潘连德,梁天红,等.松江鲈鱼(Trachidermus f asciatus)胚胎发育的初步观察[J].复旦大学学报(自然科学版),2004,43(2):250-254.
[2]Mcpherron AC,Lawler AM,Lee SJ.Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily membe[J].Nature,1997,387:83-90.
[3]赵浩斌,彭扣,王玉凤,等.鱼类肌肉生长抑制素研究进展[J].水生生物学报,2006,30(2):227-231.
[4]Maccatrozzo L,Bargelloni L,Radaelli G,et al.Characterization of the myostatin gene in the gilthead seabream(Sparus aurata):sequence,genomic structure,and expression pattern[J].Mar Biotechnol(NY),2001,3(3):224-230.
[5]Garikipati DK,Gahr SA,Roalson EH,et al.Characterization of Rainbow Trout Myostatin-2 Genes(rtMSTN-2a and-2b):Genomic Organization,Differential Expression,and Pseudogenization[J].Endocrinology,2007,148(5):2106-2115.
[6]卢祥云,张营,王星果,等.松江鲈肌肉生长抑制素基因克隆和序列特征分析[J].动物学研究,2010,31(4):387-394.
[7]郭新红,刘少军,刘巧,等.鱼类线粒体DNA研究新进展[J].遗传学报,2004,31(9):983-1000.
[8]Maccatrozzo L,Bargelloni L,Cardazzo B,et al.A novel second myostatin gene is present in teleost fish[J].FEBS Lett,2001,509(1):36-40.
[9]Rodgers BD,Weber GM.Sequence conservation among fish myostatin orthologues and the characterization of two additional cDNA clones from Morone saxatilis and Morone americana[J].Comp Biochem Physiol B Biochem Mol Biol,2001,129(2-3):597-603.
[10]Rodgers BD,Garikipati DK.Clinical,agricultural,and evolutionary biology of myostatin:a comparative review[J].Endocr Rev,2008,29(5):513-534.
[11]De la Rosa-Reyna XF,Rodríguez Pérez MA,Sifuentes-Rincón AM.Microsatellite polymorphism in intron 1 of the bovine myostatin gene[J].J Appl Genet,2006,47(1):55-57.
[12]姜运良,李宁,赵兴波,等.猪肌肉生长抑制素基因侧翼区新微卫星标记的鉴定及分析[J].遗传学报,2004,31(5):480-484.
[13]Maccatrozzo L,Bargelloni L,Patarnello P,et al.Characterization of the myostatin gene and a linked microsatellite marker in shi drum(Umbrina cirrosa,Sciaenidae)[J].Aquaculture,2002,205(122):49-60.
[14]薛良义,孙升,肖章奎,等.大黄鱼肌肉生长抑制素基因微卫星序列多态性分析[J].中国生物化学与分子生物学报,2008,24(10):980-985.
