气候变化驱动下树种迁移过程初探
2011-01-26刘晓梅布仁仓
刘晓梅 ,布仁仓 ,郭 锐
(1.中国科学院沈阳应用生态研究所,辽宁 沈阳 110016;2.中国科学院研究生院,北京 100039)
近百年来,地球气候正经历着一次以全球变暖为主要特征的显著变化,平均地表温度将增加2~4.5℃、极端降雨事件发生频率增加、植物生长季延长等[1]。气候变化导致的生态环境的变化越来越受到人们的关注,作为陆地生态系统重要组分的森林生态系统,对未来气候变化的响应更是人们关注的重点[2]。气候是森林生态系统功能重要的驱动力,其变化会改变森林生态系统的特征和功能[3]。近百年气候变化的速度超过了以往的变化速度,为了适应新的温度及降雨格局,树种将逐步向适于生存的环境迁移。研究表明过去森林在极端气候条件下,以每个世纪超过50 km的速率进行迁移[4]。在气候变化驱动下,树种通过改变其物候或生理反应来适应气候变化,或迁移到更适合的生境,甚至是灭亡[5]。在气候变化中存活下来的物种可能具有高的基因多样性、广的生态幅、可塑性高的表型、后代时间短及长距离种子扩散适应等[6]。如果不具备这些特征,树种就可能灭亡。可见,在气候变化驱动下树种迁移过程的研究显得极为重要。
树种在迁移过程中,可能遇到前所未遇的干扰,如当地树种的竞争、森林采伐[7]、病虫害及生境破碎化,这些可能改变迁移速率。目前的研究主要集中在森林树种组成及分布范围:(1)分析树种在气候变化条件下潜在分布的变化[8],注重分布格局的研究,而忽略过程对格局的影响;(2)在林窗、景观尺度上研究树种的潜在分布[9],无法回答研究区外的树种迁移和分布格局,过高估计树种灭亡;(3)气候变化预案中存在很大的不确定性[10]。(4)大部分模型研究基于气候与树种的一种平衡关系,很少或者没有考虑物种的耐性、迁移能力、迁移速率以及迁移障碍等因素的影响[11],并且缺乏对极端气候事件及季节变化的考虑等。本文根据种子生产、种子扩散、种子库、种子萌发、幼苗建成及幼树长成5个阶段的特征及影响因素,从机理上研究整个迁移过程。
1 树种迁移过程
1.1 种子生产
种子是重要的繁殖器官,通过对生态环境不断适应植物形成了不同的繁殖策略。种子生产为树种迁移过程提供种源,许多植物种群受种子限制,即使是多年生植物种群也不例外[12]。捕食者饱食假说认为丰富的种源可以增加种子逃脱捕食者及建群的可能性。从群落的角度看,种子缺乏会对处于演替初期的群落及较低持久力的土壤种子库产生很大的影响,可以改变植物群落的组成、限制局部植物的分布及群落的结构,甚至可能导致入侵物种占有主导地位[13],这样,入侵种占据空间,阻碍树种迁移的过程,导致树种有序迁移过程的中断,甚至导致树种死亡。
种子大小及重量对种子扩散、萌发、定居成功率、幼苗大小及生长速度均有影响,并与种子埋藏的垂直高度相关,决定种子库的动态[14]。种子大小对于树种迁移过程各有利弊,种子大,储存的营养多,发芽率高、幼苗生长速度快,生物量积累水平高,受环境的影响小,但目标大,不易被埋藏,易遭受取食。而种子小,更容易扩散,实现长距离迁移,有相对较高的生长率[15]。种子的翅、冠毛、刺、芒和毛等附属物可以辅助实现种子扩散。例如在不可预测的大尺度干扰的生境,植物区系中会存在很多能产生大量风媒种子的物种,借助风媒种子繁殖更新能使植物适应空间上不可预见的干扰[16]。
气候变化会影响种子生产,CO2浓度升高,种子产量会增加[17],种子产量增加伴随着质量降低的现象在木本植物中少见[18]。所以,气候变化背景下,树种可能有足够的种源,从而有利于其迁移过程。高山环境中,海拔高的树种在有利的气候条件下能成产生高品质的种子,气候变暖条件下,树种可以实现向高海拔迁移,使山顶植物多样性增加[19]。而种子的其他特征(形状、附属物)等,在较长的时间内,也可能发生改变来适应气候变化。
1.2 种子扩散
自20世纪70年代,种子的扩散方式和扩散过程就被植物学家和鸟类学家等广泛关注,现在已成为进化生态学研究的重要内容之一。种子扩散是树种迁移过程的核心,主要从种子扩散方式及扩散阶段两方面进行探讨。
1.2.1 种子扩散方式 种子扩散方式依据扩散载体可以划分为风扩散、动物扩散及水扩散3种。风扩散具有较强的季节性。适应风力扩散的种子一般细小而轻,能悬浮在空气中被风力吹送到远处,其次是果实或种子的表面有絮毛、果翅,或其它有助于承受风力的特殊结构。对于冠层树种、攀缘树种及开阔地的先锋树种来说,风扩散是普遍存在的。作为气候因子的风,对树种迁移过程影响最为直接和迅速,并且能实现长距离扩散,这对树种迁移过程很重要。
动物既是种子的捕食者,同时也是散播种子的重要的媒介,植物与动物的不同组合而形成特殊体系,体现了一种互生现象[20],在促进种子扩散方面有重要的作用:首先是扩散作用,食果动物通过它们的行为及生境的偏好选择有利于种子发芽的生境散播种子[21]。其次,动物的消化系统对种子的萌发起到了一定的作用,软化种皮,增加对水和空气的通透性,剥离种子果肉,这些都有利于种子发芽[22]。另外,通过消化系统的种子周围的排泄物可以增加种子有效定居的营养有效性。当然动物也对种子扩散产生负面的影响:捕食减少了种子数量,可能产生种子限制,并且捕食降低了种子的发芽率[23]。在气候变化背景下,植物和动物都会受到影响,例如植物物候期的改变,动物发生迁徙,或迁徙时间、距离的改变等,都会影响到动物与植物的互惠或者协同进化关系,甚至打破这种关系,建立新的关系,从而影响树种迁移过程。
水流也是种子扩散的载体之一,相关研究较少。降水量的变化影响河流、湖泊等水体,从而影响以水流为载体的种子扩散,依赖漂流可以进行长距离种子扩散[24]。
人类活动是森林生态系统动态的重要驱动力,也影响着种子扩散:人类运输农作物,或者无意识的将种子带到新的生境都可以实现种子的迁移[24]。采摘果实,捕杀动物等会打破种子扩散链,影响扩散过程。在气候变化下,人类可以发挥能动意识,打破生境破碎化对树种迁移的限制,人为调控树种迁移。
总之,种子扩散过程不是局限在一种扩散方式,而可能是多种扩散方式综合作用,并且扩散方式具有一定的延展性。那么,在气候变化驱动下,树种原来的扩散方式是否会发生改变来促进树种的迁移,这需要一定的时间尺度,需要我们更深入的探讨。气候变化会影响扩散载体,一方面直接影响扩散载体,例如鸟类迁徙时间的改变、扩散距离的缩短等,并且扩散载体可能给树种造成致命的伤害,例如飓风,洪水、动物取食等。另一方面,树种对气候变化的响应来间接影响扩散载体,例如种子的产量,从而影响整个种子扩散过程。
1.2.2 种子扩散的阶段性 种子扩散是一个复杂、多阶段的过程,一般分为初级扩散和次级扩散,大部分研究强调种子的初级扩散,而忽略了次级扩散,而次级扩散在植被再生产过程中是一个重要的、不容忽视的过程。每个扩散阶段的作用是不同的,初级扩散主要为了逃脱捕食者和新斑块的扩张,而次级扩散主要是逃脱捕食者和定向扩散,通过定向扩散,可以将种子扩散到适宜幼苗更新的生境。定向扩散最重要的表现是对种子埋藏,埋藏的种子比起地表面的种子具有较低的被非挖穴动物带走的几率[25],可以逃避一些恶劣的环境,并且比起地表面的种子更有可能萌发。
气候变化条件下,一方面,种子扩散阶段可能会增加,将会影响树种的迁移,例如初级扩散后,种子可能进行次级扩散,将种子扩散到更远的适宜生境,这关键取决于扩散载体(动物)生活习性的改变等,另一方面,种子扩散的阶段性是否会被中断,从而影响树种迁移。
1.3 种子库
种子成熟后,最终会到达地面或者埋在土壤里,形成种子库。作为植物定居过程中的一种优势策略,能很好地调节种子雨的时空变化,减少种子和幼苗的年变化,在空间、时间上不可预测干扰中发挥重要作用。土壤种子库具有不同的时空格局及动态(图 1)。

图1 土壤种子库动态总模型(引自Simpson)
在气候变化背景下,种子库是树种迁移过程另外的一个种源,尤其是树种迁移的边缘地带,种子库累积种子,当环境条件适宜时,立即萌发,促进树种的迁移。并且,树种迁移过程形成的种子库可以通过休眠调节种子的萌发时间,度过不良环境或对付不可知的干扰因素,从而实现成功迁移。当然种子休眠也有不利的一面,在最易被取食阶段,可能遭受高比例的取食,使其成为幼苗的几率变小。
1.4 种子萌发、幼苗建成及幼树长成成树
在许多生态系统中,种子萌发和幼苗定居是物种生活史中主要的瓶颈,比种子产量本身对群落动态和扩张的限制作用更大[26]。近来研究表明,在高纬度地区植被分布区改变及较大尺度的植被变化,定居可能是比扩散更为关键[27]。可见,种子扩散后续阶段的作用也是不容忽视的。种子萌发是植物进入营养生长阶段的关键一步,除自身的萌发特性以外,光照、温度、湿度以及地被物苔藓、草等条件都是影响种子萌发的重要因子。幼苗是植物生活史中对环境条件反映最为敏感的时期,其定居因素除了外界环境因素影响外,还受到密度(竞争)制约影响。种子扩散到达新生境,开始可能以种间密度制约(竞争)为主,随着树种的大量迁入,种内、种间密度制约会越来越重要。
气候变暖对于幼苗定居的影响存在争议,一些研究表明温度增加会减少幼苗的出现和定居。气候变暖成为种子萌发和早期幼苗的存活的限制因子,而降雨量的增加可以减缓这种限制作用[28]。气候变化对幼苗定居的影响因不同阶段、对其他因子的依赖性及不同的物种而不同。基于现行气候条件下相关研究,树种迁移过程研究应充分考虑气候变化驱动下外界环境因素及生物因素(竞争等)对种子萌发及幼苗建成的影响。
2 相关模型
由于扩散阶段的复杂性及时间、空间确定种子命运难度大,大部分野外试验局限在小时空尺度的一个或几个阶段,目前定性模型有种子命运模型、补充限制模型及时空模型[29]。Nathan的时空模型是基于种子扩散的空间形式总结的模型。强调种子扩散的时空变化对植物种群和群落的空间结构的影响,对树种迁移过程的研究有很好的借鉴意义(图2)。
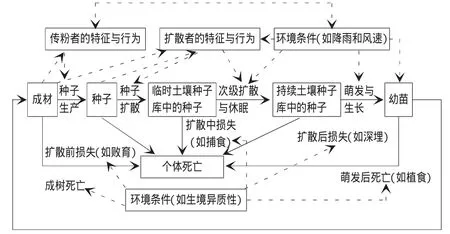
图2 时空模型(引自Nathan)
气候变化和树种迁移是一个几十年甚至上百年的过程。不论是时间尺度还是空间尺度都超过了人类现有的实验能力。应用模型,可以用数学和逻辑的方法精确定义树种的生物学特性、物种扩散等来回答试验科学无法回答的问题。在气候变化驱动下,大时空尺度的植被响应模型可以归并为生物地理相关模型、生态响应面模型、立地模型、植物生理模型以及统计模型等五类[30]。各种模型正在逐步融合,而在区域景观尺度上的建模引起广泛的关注,空间直观景观模型已成为研究气候变化下森林生态系统动态的重要工具,模拟较大时空尺度的森林更新演替过程[31]。Landis7.0 Pro是空间直观景观模型的典型代表,在Landis 6.0 pro的基础上加以改进,考虑了树种的结实性、扩散距离、耐阴性及立地相对密度等因素,更加接近真实的生态学过程,更好地模拟气候变化驱动下整个大时空尺度的森林景观动态。
3 结语
本文从一个整体、机理的新角度来探讨气候变化驱动下树种迁移过程。这方面研究很少,却极其重要。大部分研究局限在两个方面:一方面是现行气候条件下小时空尺度一个或几个阶段的研究;另一方面是气候变化背景下对大时空尺度的森林分布、组成、演替、生产力、森林病虫害、火灾等的影响,而对产生这些影响的原因缺乏机理性的解释。树种迁移过程是综合小时空尺度的研究来实现对大尺度森林生态系统响应的机理性的解释,从整体上分析了树种的迁移过程。
明确树种迁移过程的关键阶段及影响因素,可以为人为促进迁移提供理论支持,为气候变化下森林资源的可持续利用及国家林业发展战略提供依据。总之,树种迁移过程是一个大时空尺度、复杂的过程,今后需要进行更深入的研究。
树种迁移过程是一个较大时空尺度的生态过程,传统的方法不能满足要求,模型的研究必将成为一种趋势,区域景观尺度的模型研究成为焦点,各类定量模型正在逐步融合,从整体上探究树种的迁移过程,为大时空尺度的模型的研究及优化提供理论支持。
[1]IPCC.Climate change 2007:The physical science basis.[M].Cambridge:Cambridge University Press.2007:996.
[2]刘国华,傅伯杰.全球气候变化对森林生态系统的影响[J].自然资源学报,2001,16(1):71-78.
[3]Woodall C,Oswalt C,Westfall J,et al.An indicator of tree migration in forests of the eastern United States[J].Forest Ecology and Management,2009,257(5):1434-1444.
[4]Schwartz M.Modeling effects of habit fragmentation on the ability of tree to respond to climate warming[J].Biodiversity and Conservation,1992,2:51-61.
[5]Davis M,Shaw R.Range shifts and adaptive responses to quaternary climate change[J].Science,2001,292:673-679.
[6]Vitt P,Havens K,Kramer A,et al.Assisted migration of plants:Changes in latitudes,changes in attitudes[J].Biological Conservation,2010,143(1):18-27.
[7]Bu R,He H,Hu Y,et al.Using the LANDIS model to evaluate forest harvesting and planting strategies under possible warming climates in Northeastern China[J].Forest Ecology and Management,2008,254(3):407-419.
[8]冷文芳,贺红士,布仁仓,等.东北落叶松属植物潜在分布对气候变化的响应[J].辽宁工程技术大学学报,2007,26(2):289-292.
[9]延晓东,赵士洞.气候变化背景下小兴安岭天然林的模拟研究[J].自然资源学报,1999,14(4):
[10]Xu C,Gertner G,Scheller R.Uncertainties in the response of a forest landscape to global climatic change[J].Global Change Biology,2009,15(1):116-131.
[11]刘国华,傅伯杰.全球气候变化对森林生态系统的影响[J].自然资源学报,2001,16(1):71-78.
[12]Jacquemyn H,Brys R,Jongejans E.Seed limitation restricts population growth in shaded populations of a perennial woodland orchid[J].Ecology,2010,91(1):119-129.
[13]Seabloom E,Harpole W,Reichman Q,et al.Invasion,competitive do minance,and resource use by exotic and native California grassland species[J].Proceedings of the National Academy of Science USA,2003,100:13384-13389.
[14]Bekker R,Bakker J,Grandin U,et al.Seed size,shape and vertical distribution in the soil:indicators of seed longevity[J].Functional Ecology,1998,12(5):834-842.
[15]He Y,Wang M,Wen S,et al.Seed size effect on seedling growth under different light conditions in the clonal herb Ligularia virgaurea in Qinghai-Tibet Plateau[J].Acta Ecologica Sinica,2007,27(8):3091-3108.
[16]刘志民,蒋德明,高红瑛.植物生活史繁殖对策与干扰关系的研究[J].应用生态学报,2003,14(3):418-422.
[17]Ladeau S,Clark J.Rising CO2levels and the fecundity of forest trees[J].Science,2001,292:95-98.
[18]Way D,Ladeau S,Mccarthy H,et al.Greater seed production in elevated CO2is not accompanied by reduced seed quality[J].Global Change Biology,2010,16(3):1046-1056.
[19]Jackson S,Sax D.Balancing biodiversity in a changing environment:extinction debt,immigration credit and species turnover[J].Trends in Ecology&Evolution,2010,25(3):153-160.
[20]Hubbell S.Seed predation and the coexitence of tree species in tropical forests[J].Oikos,1980,35:214-229.
[21]Valido A,Nogales M.Frugivory and seed dispersal by the lizard Gallotia galloti(Lacertidae)in a xeric habitat of the Canary Islands[J].oikos,1994,70(3):403-411.
[22]Traveset A,Verd M,Levey D,et al.A meta-analysis of the effect of gut treatment on seed germination[J].CABI Publishing,2002,339-350.
[23]彭闪江.植物更新过程中种子和幼苗死亡的影响因素[J].广西植物,2004,24(2):113-121.
[24]Nathan R,Schurr F,Spiegel O,et al.Mechanisms of long-distance seed dispersal[J].Trends in Ecology&Evolution,2008,23(11):638-647.
[25]Cahalan V.Caching and recovery of food by the western fox squirrels[J].Journal of Wildlife management,1942,6:338-352.
[26]Korner C,Alpine plant life:functional plant ecology of high mountain ecosystems[M].Springer Verlag.2003,291-297.
[27]Alsos I,Eidesen P,Ehrich D,et al.Frequent long-distance plant colonization in the changing Arctic[J].Science,2007,316(5 831):1606.
[28]Shevtsonva A,Graae B,Jochum T,et al.Critical periods for impact of climate warming on early seedling establishment in subarctic tundra[J].Global Change Biology,2009,15(11):2662-2680.
[29]肖治术,张志彬,王玉山.以种子为繁殖体的植物更新模型研究[J].生态学报,2003,22(4):70-75.
[30]李双成.植物响应气候变化模型模拟研究进展[J].地理科学进展,2001,20(3):217-225.
[31]徐崇刚,胡远满,常 禹,等.空间直观景观模型LANDISⅠ.运行机制[J].应用生态学报,2004,15(5):837-844.
