拟南芥耐硒突变体的筛选和鉴定
2011-01-23慈凌坤狄东伟卢云峰曹树青
慈凌坤, 江 力, 狄东伟, 张 军, 卢云峰, 曹树青
(合肥工业大学生物与食品工程学院,安徽合肥 230009)
硒是环境中一种重要的生命元素,随着人类对硒生化功能认识的日渐深入,促进了植物硒的生理机能研究的进一步发展。硒在植物体内以无机硒和有机硒2种形式存在,大部分为有机硒形式,约占总量的80%以上,主要是硒代胱氨酸、硒代半胱氨酸和硒代蛋氨酸,硒以硒代含硫氨基酸形式直接参与蛋白质合成,是重要的抗氧化酶类谷胱甘肽过氧化物酶(GSH-Px)的组成部分[1]。硒在植物上的生理功能涉及到生长、发育、营养、代谢、产量、品质和抗性等许多方面[2-3],硒发挥这些功能的作用机制仍然不完全清楚。
拟南芥是重要模式植物,由于近年来基因组学及遗传学的发展,它的完整基因组已被成功测定,因此为新基因、基因功能方面的研究提供很多优势。本实验采用不同质量浓度的亚硒酸钠(Na2 SeO3)处理野生型和突变体拟南芥,通过对其形态学及生理生化指标的测定来筛选并鉴定耐硒突变体。本研究不仅为耐硒基因克隆及功能分析奠定基础,而且对于揭示植物耐硒的分子机理也具有重要意义。
1 材料与方法
1.1 材料与培养
本实验所用拟南芥(Arabidopsis)生态型为Colum bia,野生型(W T)和突变体(Mutant)来自美国拟南芥种质资源中心。
实验采用2种方法培养植株:①培养基培养[4]:种子经0.1%HgCl2消毒3 min后,用无菌水冲洗4~5次,将种子点种在1.5%蔗糖和0.8%琼脂的培养基中;并用封口膜封装,于4℃条件下处理2 d后转至光照为l00μmol/(m2·s2)、温度为22~23℃的培养室中。②土壤培养:种子播种于装有土壤的盆钵中,置于光周期为光照16 h,黑暗8 h、光照为100 μmol/(m2·s2)、温度为 22~ 23 ℃的培养室内,播种和开花时各浇1次营养液,并根据植株生长需要浇以适量水。
1.2 筛 选
[5-7]和预试验,确定 50 mg/L Na2 SeO3为筛选质量浓度。普通MS培养基上点种,育苗7 d后移入50 m g/L Na2SeO3选择培养基,以根长作为筛选指标,选择根长明显长的作为候选突变体。
1.3 鉴 定
1.3.1 正常培养条件下的生长状况
将野生型和突变体的种子直接播种于土壤和普通培养基中,不进行任何处理,定期观察拍照,比较突变体和野生型的生长状况。
1.3.2 不同质量浓度Na2 SeO3处理下的表型
设置 6 个质量浓度(50、80、100、150、300、500 mg/L)Na2 SeO3和1个对照,对生长 21 d的拟南芥野生型和突变体叶片进行涂抹,48 h后观察不同质量浓度下野生型和突变体的生长情况,拍照记录并分析。
1.3.3 不同质量浓度Na2 SeO3处理下的发芽率
设置 5 个质量浓度(10 、20 、30 、40 、50 mg/L)Na2SeO3和1个对照(0 m g/L Na2SeO3的 MS培养基),分别将野生型和突变体种子点种在含有不同质量浓度Na2 SeO3的培养基中,成5×10矩阵,封口后放入4℃冰箱中春化2 d后置于培养室中培养,观察记录发芽率,直至第4天种子的发芽率不再变化为止。
1.3.4 蛋白质量浓度和酶活性的测定
设置2个Na2 SeO3质量浓度(50、80mg/L)和1个对照,将生长21 d的拟南芥叶片间隔24 h涂抹2次后,72 h后分别测定突变体和野生型叶片可溶性蛋白质量浓度[8]和GSH-Px活性[9]。
2 结果与分析
2.1 筛 选
突变体的筛选如图1所示,箭头所示为根长明显较长的植株,挑选出来移栽到土壤培养基中作为候选突变体培养。

图1 突变体的筛选
2.2 候选突变体的土壤培养
候选突变体的整个生长周期如图2所示。由于基因突变的复杂性,移栽到土壤培养基中的突变体,只有一部分能够正常生长,经过一段时间的培养,最后有2株候选突变体成活,命名为vse1和vse2。

图2 候选突变体的生长周期
2.3 野生型和突变体在正常生长条件下的表型
正常生长条件下,突变体和野生型的表型如图3所示,由图3可以看出,两者的表型没有明显的区别。
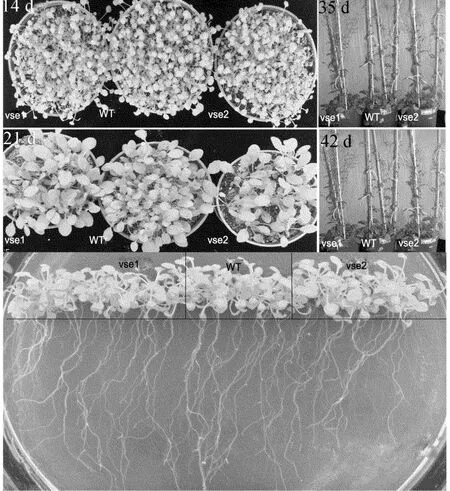
图3 野生型和突变体在正常生长条件下的表型
图3下方是正常培养基中野生型和突变体培养28 d后的比较,显示普通培养基中,突变体vse1、vse2与野生型没有明显的区别。
2.4 野生型和突变体在硒盐胁迫下的表型
野生型和突变体vse1、vse2在不同质量浓度Na2 SeO3处理下的表型比较如图4所示。从图4可看出,50mg/L Na2 SeO3处理后,突变体 vse1、vse2与野生型无明显差异;80 m g/L Na2SeO3处理后,野生型的长势明显较突变体vse1、vse2差,且有部分枯萎;100 mg/L Na2 SeO3处理后,野生型基本枯萎,突变体vse1的长势较好;150mg/L Na2 SeO3处理后,野生型和2个突变体大部分都已经枯萎;300、500 mg/L Na2SeO3处理后,野生型和突变体全部枯萎。说明在 50~100 mg/L Na2 SeO3胁迫下,突变体vse1和vse2比野生型有较好的耐受性。
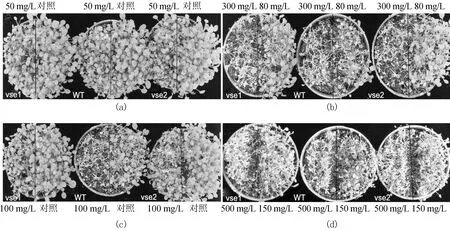
图4 野生型和突变体在硒盐胁迫下的表型
2.5 野生型和突变体在硒盐胁迫下的发芽率
野生型和突变体在硒盐胁迫下的发芽率如图5所示。
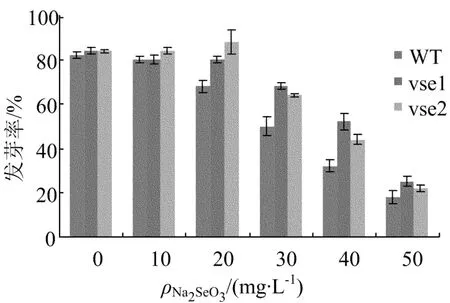
图5 野生型和突变体在硒盐胁迫下的发芽率
从图5可看出,普通MS培养基中,突变体vse1、vse2与野生型发芽率无差异;10 mg/L Na2 SeO3胁迫下,突变体 vse1、vse2与野生型发芽率无明显差异;20、30、40 m g/L Na2SeO3胁迫下,野生型发芽率较突变体vse1、vse2差异很大;30 mg/L Na2SeO3胁迫下,突变体 vse1、vse2发芽率分别比野生型高出 18%、14%;40 mg/L Na2 SeO3胁迫下,突变体 vse1、vse2发芽率分别比野生型高出20%、12%;50 mg/L Na2 SeO3是筛选的临界质量浓度,突变体vse1、vse2发芽率高于野生型。
2.6 野生型和突变体在硒盐胁迫下蛋白质量比
野生型和突变体在硒盐胁迫下可溶性蛋白质量比的变化如图6所示,与对照组相比,80mg/L Na2SeO3处理后,拟南芥突变体幼苗的可溶性蛋白质量比明显增加,vse1和vse2可溶性蛋白质量比分别较对照组增加125%和188.9%,W T可溶性蛋白质量比变化不明显。这说明硒盐胁迫下,突变体通过增加可溶性蛋白的质量比增强了自身耐胁迫能力。

图6 不同质量浓度Na2 SeO 3处理下可溶性蛋白质量比
2.7 野生型和突变体在硒盐胁迫下酶活性
野生型和突变体在硒盐胁迫下GSH-Px活性的变化如图7所示,野生型和突变体的GSHPx活性均随着Na2 SeO3质量浓度的增加而呈现递增趋势,但野生型的增长趋势小于突变体。80mg/L Na2 SeO3胁迫下,突变体 vse1、vse2 GSH-Px活性分别比对照增加 280%和219%,WT比对照增加134%。这说明硒胁迫下,耐硒突变体的GSH-Px的酶活性明显增强,且这种趋势比野生型更加明显。
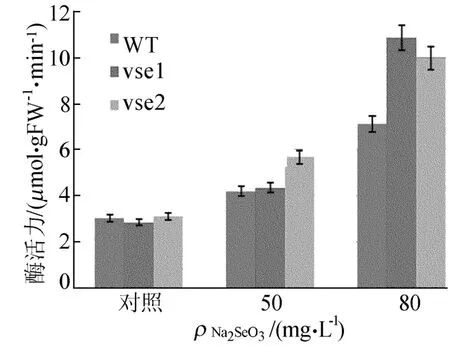
图7 不同质量浓度Na2SeO3处理下GSH-Px活性的变化
3 讨 论
突变体筛选方法是很重要的,越是专一的筛选条件,就越有机会直接获得所期望的突变体。可以采用在外源硒作用下拟南芥的根弯曲试验和发芽试验,本实验采取以根长为指标的筛选方法。实验中采用的拟南芥突变体库是利用一个化学诱导激活启动子/增强子的 T-DNA载体(XVE载体)构建成的。由于T-DNA标记或相邻的基因可被XVE系统诱导激活,或被T-DNA破坏导致功能缺失,该突变体库可以用于大规模筛选鉴定功能缺失性和功能获得性突变体[10]。
有研究显示,较低质量浓度硒(50 mg/L)有利于蛋白质的合成,微量元素硒对高等植物中GSH-Px活性影响是非常明显的[9]。本实验中,经过80 mg/L N a2SeO3处理后,vse1和vse2叶片中的可溶性蛋白含量大幅增加,野生型、突变体vse1和vse2的酶活性分别增加了134%、280%和219%。
耐硒植物耐受高质量浓度硒的机理在于它能够减少硒代半胱氨酸和硒代蛋氨酸在细胞内的质量浓度,这些氨基酸掺入到蛋白质后可以对植物蛋白的结构、肽键状态产生影响,从而导致蛋白功能的损害,如果硒被转化为非蛋白硒代氨基酸,诸如硒-甲基硒代半胱氨酸或硒代胱硫醚及二肽等,就可以减少硒的掺入[11-12]。本实验表明,在硒盐胁迫下,突变体vse1和vse2的可溶性蛋白质量比和GSH-Px的活性较野生型明显提高,这与vse1和vse2耐硒能力较野生型强结果一致。但vse1和vse2以何种信号途径调节其耐硒以及耐硒的分子机理有待进一步研究。
[参 考 文 献]
[1]徐辉碧.硒的化学、生物化学及其在生命科学中的应用[M].武汉:华中理工大学出版社,1994:190-192.
[2]李应生,李亚男,陈大清.硒的生物学功能及植物的富硒机理[J].湖北农学院学报,2003,23(6):476-480.
[3]罗世炜,张孟琴,吴永尧.植物硒的研究与利用[J].安徽农业科学,2007,35(14):4087-4088.
[4]M urashige T,Skoog F.A revised medium for rapid grow th and bioassays w ith tobacco tissue cu lture[J].Physiolgogia Plantarum,1962,15(3):473-497.
[5]Zhang Lihong,Ackley A R,Pilon-Smits E A H.Variation in selenium tolerance and accumulation among 19 Arabidopsis thaliana accessions[J].Jou rnal of Plant Physiology,2007,164:327-336.
[6]Tagmount A,Berken A,Terry N,et al.An essential role of S-adenosyl-l-m ethionine:l-methionine S-methyltransferase in selenium volatilization by plants.Methylation of selenomethionine to selenium-methyl-l-selenium-methionine,the precursor of volatile selenium[J].Plan t Physiology,2002,130(2):847-856.
[7]Shibagaki N,Rose A,M cDermott JP,et al.Selenate-resistan tmutants of Arabidopsis thaliana identify Su ltr 1;2,a sulfate transporter required for efficient transport of sulfate into roots[J].Plan t Journal,2002,29(4):475-486.
[8]Bradford M M.A rapid and sensitivemethod for the quantitation ofmicrog ram quantitiesof protein u tilizing the principle of protein-dye binding[J].Anal Biochem,1976,72:248-254.
[9]周毅峰,吴永尧,唐巧玉.硒对大豆叶片谷胱甘肽过氧化物酶和过氧化氢酶歧化酶活性的影响[J].湖北民族学院学报:自然科学版,2005,23(2):127-129.
[10]张 健,徐金相,孔英珍,等.化学诱导激活型拟南芥突变体库的构建及分析[J].遗传学报,2005,32(10):1082-1088.
[11]陈大清.植物硒同化的研究进展及其耐硒突变体的筛选[J].氨基酸和生物资源,2004,26(2):65-70.
[12]何桂平,张从合,陈金节,等.杂交油菜及其亲本对不同胁迫的响应[J].合肥工业大学学报:自然科学版,2010,33(8):1241-1244.
