耳蜗中L-精氨酸对Ca2+-ATP酶抑制剂的拮抗作用*
2011-01-23贾学斌李兴启张瀛朱耀国
贾学斌 李兴启 张瀛 朱耀国
Ca2+在体内有非常广泛的生理作用[1~3],Ca2+参与耳蜗声机械-电转换、内耳声感受的频率选择及基底膜振动的非对称性等过程。维持细胞内Ca2+平衡是发挥正常耳蜗功能的保证。细胞内Ca2+稳态的调节机制主要包括两方面,即增加胞浆内Ca2+浓度和减少胞浆内Ca2+浓度。增加胞浆内Ca2+浓度途径有:细胞膜上的Ca2+通道开放、Na+/ Ca2+交换、Ca2+/H+交换及Ca2+被动扩散入胞等;而减少胞浆内Ca2+浓度的方式主要有Ca2+-ATP酶将Ca2+泵出细胞或泵入Ca2+库和Na+/ Ca2+交换。若Ca2+-ATP酶的功能被抑制必将引起胞浆内Ca2+超载,可导致细胞损伤甚至死亡。L-精氨酸被认为是合成NO的底物,它可在一氧化氮合酶(NOS)及黄素腺嘌呤二核苷酸(FAD)、黄素腺嘌呤单核苷酸(FMN)、四氢生物喋呤(BH4)、还原型尼克酰胺嘌呤二核苷酸(NADPH)等转氢、转电子体的协同作用下生成NO和L-胍氨酸。由此,L-精氨酸激活NO/cGMP通路,产生cGMP,进而激活磷酸激酶cGK-1对功能酶磷酸化。L-精氨酸激活NO/cGMP通路使磷酸激酶cGK-1活化后,可对多种蛋白质进行磷酸化,其中,对细胞内磷脂酶C的磷酸化将减少磷酸肌醇IP3的含量,进而起到降低胞内Ca2+浓度的作用,从而调节细胞胞浆内Ca2+的平衡。
在耳蜗中存在着NO/cGMP通路的各关键酶及Ca2+ATP酶。本实验用Ca2+-ATP酶抑制剂造模,使胞内Ca2+超载,然后加入L-精氨酸以激活NO/cGMP通路,观察耳蜗中L-精氨酸是否对Ca2+-ATP酶抑制剂有一定的拮抗作用。
1 材料与方法
1.1主要试剂和仪器 基础液:人工外淋巴液(APL)(mM):NaCl 137,KCl 5,CaCl2,NaH2PO41,Glucose 11,NaHCO312,MgCl21,用前将pH调至7.2~7.4,用全自动冰点渗透压仪调节渗透压到300±2 mOsm/L。实验试剂:L-精氨酸、环磷酸鸟苷(cGMP)、 Ca2+-ATP酶抑制剂(Cyclopiazonic acid)、一氧化氮合酶(NOS)抑制剂L-NNA、亚甲基蓝(Methylene blue)。
微量注射泵WZS-50F, STORZ冷光源, OLYMPUS解剖显微镜,数字温度计,微操纵器,电屏蔽实验台,动物人工呼吸机, TDT系统自制声控装置,ER-10C型耳塞式耳机。
1.2实验动物及分组 选用耳廓反射灵敏、体重250~350 g的健康杂色豚鼠70只,雌雄不限,随机分为7组,每组10只:①人工外淋巴液组;②Ca2+-ATP酶抑制剂组;③L-精氨酸组;④Ca2+-ATP酶抑制剂+ L-精氨酸组;⑤Ca2+-ATP酶抑制剂+环磷酸鸟苷(cGMP)组;⑥Ca2+-ATP酶抑制剂+ L-精氨酸+ 非选择性一氧化氮合酶(NOS)抑制剂组;⑦Ca2+-ATP酶抑制剂+ L-精氨酸+可溶性环磷酸鸟苷合酶抑制剂(sGC抑制剂)组。所有动物均无耳毒性药物使用史及中耳炎病史,在无噪声环境下饲养3~5天后进行实验。所有动物均以右耳为实验耳。
1.3全耳蜗灌流方法 各组动物分别全耳蜗灌流以上各药物120分钟。用5%异戊巴比妥钠(100mg/kg)经腹腔注射麻醉后,固定于电屏蔽的操作台(美国TMC公司),用数字式温度计监测体温,热水袋保持动物体温在37~39.2℃之间。行气管切开,插入气管插管行人工呼吸,呼吸频率为60次/分,潮气量3毫升/次。腹腔注射氯化琥珀胆碱(15 mg/kg),消除肌电干扰。皮下注射阿托品(15 mg/kg),减少分泌物,保持呼吸道通畅。经腹侧暴露右侧听泡。打开听泡后,在解剖显微镜下用自制尖针分别在耳蜗底回鼓阶和前庭阶的骨壁各钻一小孔(直径约0.2 mm)。在解剖显微镜下将拉制好的玻璃微管(尖端直径约50 μm)经微操纵器插入鼓阶适宜深度,并用医用粘胶封填小孔周围,避免渗漏。试剂从鼓阶入,前庭阶出,全耳蜗灌流速度为2 μl/min。每只动物行全耳蜗灌流2小时。各组中的部分动物在实验结束时换作镁蓝灌流,清楚显示蓝色液体由底回鼓阶进入,即刻出现蜗尖处蓝染,最后蓝色液体从另一孔前庭阶流出,各回耳蜗均被染成蓝色,说明该灌流过程中试剂到达蜗顶,该灌流方法确实可靠。
1.4耳蜗听神经复合动作电位(CAP)和微音器电位(CM)的记录 记录银制电极置于圆窗龛,参考电极置于颈部肌肉,地线接鼻。短声诱发的CAP放大后信号由TDT信号处理仪处理,CAP波形显示于计算机。选择短纯音(1 kHz,10~100 dB SPL,每5 dB一档,上升/下降时间为2 ms,平台期20 ms)诱发CM(选择1 kHz),记录100~70 dB的幅度,以100 dB SPL刺激声强度的幅值为100%,其余刺激声强度的CM幅值取其与100 dB SPL刺激声幅值的比值(%)。各组动物耳蜗打孔前后记录及灌流过程中每隔30分钟记录一次CAP阈值、CM幅度,连续记录 2小时。
1.5统计学方法 采用Stata统计软件进行单因素方差分析,两两比较。
2 结果
2.1各组灌流药物对CAP阈值的影响 灌流L-精氨酸组与灌流人工外淋巴液组CAP阈移比较差异无统计学意义(P>0.05);而灌流Ca2+-ATP酶抑制剂组相对于人工外淋巴液组CAP阈移26 dB,两者比较差异有显著统计学意义(P<0.01);灌流Ca2+-ATP酶抑制剂+ L-精氨酸组的CAP阈值比灌流Ca2+-ATP酶抑制剂组改善9 dB,两者间差异有显著统计学意义(P<0.01);而Ca2+-ATP酶抑制剂+ L-精氨酸组与灌流Ca2+-ATP酶抑制剂+cGMP组比较差异无统计学意义(P>0.05);在Ca2+-ATP酶抑制剂+ L-精氨酸基础上加入非选择性NOS抑制剂则使阈值提高了10 dB,两者比较差异有显著统计学意义(P<0.01);加NOS抑制剂组与加sGC抑制剂组间差异无统计学意义(P>0.05)(表1)。
2.2各组灌流药物对CM相对幅度的影响 100 dB SPL 1 kHz和70 dB SPL 1 kHz CM相对幅度变化不大。其中,灌流L-精氨酸组与灌流人工外淋巴液组比较差异无统计学意义(P>0.05);而灌流Ca2+-ATP酶抑制剂组相对于人工外淋巴组CM相对幅度下降了大约68%,两者比较差异有显著统计学意义(P<0.01);灌流Ca2+-ATP酶抑制剂+ L-精氨酸组的CM相对幅度比灌流Ca2+-ATP酶抑制剂组提高了大约25%,两者间差异有显著统计学意义(P<0.01);而Ca2+-ATP酶抑制剂+ L-精氨酸组与灌流Ca2+-ATP酶抑制剂+cGMP组比较差异无统计学意义(P>0.05);在Ca2+-ATP酶抑制剂+ L-精氨酸基础上加入非选择性NOS抑制剂则使CM相对幅度下降了大约22%,两者比较差异有显著统计学意义(P<0.01);加NOS抑制剂组与加sGC抑制剂组间差异无统计学意义(P>0.05)(表1)。
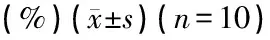
表1 各组灌流药物前后的CAP阈移(dB)及1 kHz短纯音100、70 dB SPL的CM相对幅度
3 讨论
Ca2+作为第二信使参与了机体众多的生理活动,如分泌、运动、能量代谢、细胞分裂、蛋白合成、膜通透性和离子转运等[1~3]。在耳蜗中,Ca2+与毛细胞功能的维持、机械-电转换、外毛细胞的主动收缩反应、内耳声感受的频率选择、基底膜振动的非对称性及非线性响应、毛细胞适应性以及传出神经对内耳的调控等关系密切[4,5],而维持细胞内的Ca2+浓度相对平衡是完成这些生理功能的基础。
Ca2+-ATP酶作为排出Ca2+的主要方式已受到越来越多的关注。Ca2+-ATP酶广泛存在于真核细胞中,它对细胞内Ca2+的调节是一种主动的离子转运过程,每转运一个Ca2+要消耗1~2个ATP。Ca2+-ATP酶与Ca2+有较高的亲和性,可在静息细胞内Ca2+浓度很低的情况下不断将Ca2+排出细胞或泵入胞内钙库,持续的发挥对细胞内Ca2+浓度的调节作用[1],故Ca2+-ATP酶的作用是保持相对稳定的细胞内低Ca2+。Ca2+-ATP酶分布于耳蜗中的内外毛细胞、支持细胞(如Deiters’细胞、Hensen’s细胞及柱状细胞)、血管纹的细胞、螺旋神经节细胞及耳蜗的传出传入神经纤维[6~12]。当Ca2+-ATP酶被抑制时,造成胞内Ca2+超载,对耳蜗功能的影响表现为耳蜗微音器电位(CM)幅度下降,耳蜗听神经复合动作电位(CAP)阈值提高及耳蜗内电位(EP)下降[7,13~16]。本研究结果与文献相似,可见,Ca2+-ATP酶抑制剂可使CAP阈值提高,CM相对幅度下降及非线性特性减弱。一方面是由于Ca2+超载可抑制钙库动员,使内外毛细胞、传出传入神经纤维、螺旋神经节细胞及血管纹中参与形成EP的细胞功能受到抑制;另一方面,由于Ca2+超载使供应耳蜗的血管发生痉挛,造成耳蜗细胞能量供应减少,其功能也必将受到抑制。
本实验中,在Ca2+-ATP酶抑制剂的基础上加入L-精氨酸可缓解前者对CAP阈值及CM相对幅度的影响,而加入非选择性NOS抑制剂后则L-精氨酸的作用被减弱。且加入NOS抑制剂和sGC抑制剂后对CAP阈移和CM相对幅度具有同样的作用,提示L-精氨酸在NOS作用下有改善耳蜗功能的作用,且可被非选择性NOS抑制剂和sGC抑制剂抑制。其可能的机制是:L-精氨酸是合成NO的底物,它可以活化NO/cGMP通路产生cGMP,后者可激活cGK-1,活化的cGK-1通过磷酸化过程调节各功能酶的活性,从而起到调节细胞功能的作用。因Ca2+-ATP酶抑制剂使胞内Ca2+浓度升高途径是抑制Ca2+向胞外排出和减少胞内钙库摄取Ca2+,故推测L-精氨酸对Ca2+-ATP酶抑制剂的拮抗作用是通过抑制细胞内磷脂酶C的活性,降低了磷酸肌醇IP3的含量,从而使胞内钙库减少Ca2+的释放,或者改善被Ca2+-ATP酶抑制剂抑制的Ca2+-ATP酶的活性,使向胞外排出的Ca2+及由钙库摄取的Ca2+都增多[17,18]。
本研究结果说明,Ca2+-ATP酶及NO/cGMP通路在调节细胞Ca2+平衡方面有着非常重要的作用,但其具体的作用机制尚待进一步的研究。
4 参考文献
1 Clapham DE. Calcium signaling[J]. Cell,1995,80:259.
2 Nario K,Katano I, Mori N, et al. Effect of endoplasmic Ca2+-ATPase inhibitor on cochlear potentials in the guinea pig[J]. Acta Otolaryngol,1998,118:198.
3 Lloret S, Martinez J, Moreno JJ. Influence of calcium on arachidonic acid mobilization by murine resident peritoneal macrophages[J]. Arch Biochem Biophys,1995,323:251.
4 Hudspeth AJ. How the ear's works work[J]. Nature,1989,341:397.
5 Yamoah EN, Lumpkin EA, Dumont RA, et al. Plasma membrane Ca2+-ATPase extrudes Ca2+from hair cell sterecilia[J]. J Neuroscience,1998,18:610.
6 王坚,李倩虹,陈继生. 钙与内耳毛细胞功能[J]. 生理科学进展,1992,23:145.
7 张素珍, 孙建和,赵承军,等. 豚鼠膜迷路积水模型耳蜗Ca2+-ATP酶的变化[J]. 中华耳鼻咽喉科杂志,1999,34:8.
8 Maurer J, Mann W, Baggel M. Histochemical localization of calcium ATPase in the cochlea of the guinea pig[J]. Eur Arch Otorhinolaryngol,1992,249:176.
9 Apicella S, Chen S, Bing R, et al. Plasmalemmal ATPase calcium pump localizes to inner and outer hair bundles[J]. Neuroscience,1997,79:1 145.
10 Crouch JJ, Schulte BA. Expression of plasma membrane Ca2+-ATPase in the adult and dedeveloping gerbil cochlea[J]. Hear Res,1995,92:112.
11 Ichimiya I,Adams JC,Kimura RS. Immunolocation of Na+, K+-ATPase, Ca2+-ATPase, calcium-binding proteins, and carbonic anhydrase in the guinea pig inner ear[J]. Acta Otolaryngol,1994,114:167.
12 Schulte BA. Immunohistochemical location of intracellular Ca2+-ATPase in outer hair cells, neurons and fibrocytes in the adut and developing inner ear[J]. Hear Res,1993,65:262.
13 Li WD, Zhao L, Jiang S, et al. Effects of high intensity impulse noise on ionic concentrations in cochlear endolymph of the guinea pig[J]. Chinese Med J(English),1997,110:883.
14 Ikeda K, Morizono T. Electrochemical profile for calcium ions in the stria vascularis: cellular model of calcium transport mechanism[J]. Hear Res,1989,40:111.
15 Nario K,Katano I, Mori N, et al. Effect of endoplasmic Ca2+-ATPase inhibitor on cochlear potentials in the guinea pig[J]. Acta Otolaryngol,1998,118:198.
16 Shen J,Harada N,Nakazawa H, et al.Role of nitric oxide on ATP-induced Ca2+signaling in outer hair cells of the guinea pig cochlea[J]. Brain Research, 2006,1 081:101.
17 韩济生,主编. 神经科学原理(第二版)[M]. 北京:北京医科大学出版社 ,1999.383~392.
18 Patel MR,Stamat JC,Zdanski CJ, et al.Nitric oxide in glutamate-induced compound action potential threshold shifts[J].Hearing Research, 2008,239:54.
