喀斯特森林恢复过程中优势乔木树种种间联结性分析
2010-12-05杨春玉刘绍飞喻理飞
杨春玉,刘绍飞,喻理飞
(1.贵州大学 林学院,贵州 贵阳 550025;2.贵州茂兰国家级自然保护区 管理局,贵州 荔波 558400)
种间联结指不同物种在空间分布上的相互关联性,是对物种在不同生境中相互影响相互作用所形成的有机联系的反映,对揭示种间相互作用和群落的组成及动态具有重要的意义。在贵州生境复杂的喀斯特森林区对种间联结性研究还较少,已有的研究报道[1-2]集中于喀斯特森林恢复过程中所有植物之间的种间关系的探索,而森林群落自然恢复发生于建群层乔木优势树种之间替代,针对乔木层树种种间关系开展研究,对揭示南方喀斯特森林恢复规律,指导退化森林恢复与重建有重要意义。
1 自然条件与植被概况
研究地为贵州省茂兰国家级自然保护区。25°09′20″~25°20′50″N,107°52′10″~108°05′40″E。主要出露岩石为纯质石灰岩和白云岩,属裸露型喀斯特地貌。该区年平均气温为18.3℃,有效积温5 767.9℃;年降水量为1 320.5 mm;年均相对湿度为80%;年日照时数为1 272.8 h,日照百分率29%,属中亚热带季风湿润气候。区内土壤以黑色石灰土为主,土层浅薄且不连续。原生性植被为常绿落叶阔叶混交林,现存植被包含自然恢复各阶段的群落类型。区内退化群落的自然恢复过程分为草本群落、草灌群落、灌丛灌木、乔灌过渡、乔林和顶极群落阶段等6个恢复阶段[1]。笔者选择已形成林分的乔灌过渡阶段、乔林阶段和顶极群落阶段的群落开展研究。各阶段群落特征如下:
乔灌过渡阶段群落特征:群落高度为7~12 m,向乔、灌、草本层分化,乔木层郁闭度为0.4~0.5,灌木层盖度为65%~70%,草本层盖度为15%~20%。主要木本植物有川钓樟Lindera pulcherima,圆果化香 Platvcarva longines,香叶树 Lindera communis,黄连木 Pistacia chinensis,天峨槭 Acer wangchii,鸡仔木 Sinoadina racemosa,野柿 Diospyros kaki var.sylvestris,青冈 Cyclobalanopsis glauca,朴树 Celtis sinensis,短萼海桐 Pittosporum brevicalyx,齿叶黄皮 Clausena dunniana,青篱柴 Tirpitzia sinensis,齿叶铁仔Myrsine semiserrata等;草本植物有薹草1种Carex sp.,冷水花 Pilea notata,肾蕨Nephrolepis auriculata,江南卷柏 Selaginella moellendorfii,石韦 Pyrrosia lingua,吊石苣薹 Lysionotus pauciflorus,茜草 Rubia cordiflia等。
乔林阶段群落特征:群落高度为14~18 m,乔、灌、草本层分化明显,乔木层郁闭度为0.6~0.8,灌木层盖度为45%~50%,草本层盖度为15%~20%。主要木本植物有光皮梾木Cornus wilsoniana,香叶树,野柿,翅荚香槐Cladrastis polycarpa,南酸枣Choerospondias axillaris,短萼海桐,圆果化香,天峨槭,丝栗栲Castanopsis fargesii,十大功劳Mahonia sp.,苦木Picrasma quassioides和齿叶黄皮等。草本植物有冷水花,马蓝Strobilanthes cusia,肾蕨和薹草1种,翠云草Selaginella uncinata,江南卷柏等。
顶极群落阶段群落特征:群落高度为15~20 m,具有明显的层间植物,乔木层郁闭度为0.7~0.9,灌木层盖度为45%~50%,草本层盖度为15%~20%。主要木本植物有光皮梾木,短萼海桐,天峨槭,多脉青冈yclobalanopsis multinervis,云贵鹅耳枥Carpinus pubescens,粗柄楠Phoebe crassipedicella,多穗石栎Lithocarpus plystachyus,短叶白楠Phoebe neuratha var.brevifolia,长叶木兰Magnolia paenetalauma,天仙果 Ficus erecta var.beecheyana,掌叶木 Handeliodendron bodinieri,龙须藤Bauhinia championii,巴东荚蒾Viburnum henryi,圆果化香和香叶树等。草本植物有翠云草,马蓝,冷水花,日本蛇根草Ophiorrhiza japonica和云南羊耳蒜Liparis distans等。
2 研究方法
2.1 取样方法
采用空间代替时间[3]的方法,在乔灌过渡阶段、乔林阶段和顶极阶段的森林群落中分别设置36个10 m×10 m样方,记录各样方中植株的种名、株数、树高、胸径、冠幅和生境等。
2.2 测度方法
主要研究对象为3个阶段的森林群落主林层中重要性在前10位树种。树种重要性以树种在森林群落中的重要值确定。树种重要值 =相对频度 +相对密度 +相对显著度[4]。
种间关联测定以物种在样方中出现、不出现为依据,用2×2列联表求出种间的共存概率和关联强度。统计内容包括各样地出现的物种数、物种出现的样方数及种对间的出现关系[5]。
2.2.1 总体关联性检验 根据Schluter[6]提出的基于出现一不出现数据的方差比率来检验。其计算公式为t=(T1+T2+T3+ … +TN)/N;Pi=ni/N。其中:S为总物种数,N为总样方数,Tj为样方j内出现的研究物种的总数,ni为物种i出现的样方数,t为样方中种的平均数。VR>1,种间为正的净关联;VR<1,种间为负的净关联;VR=1,则符合所有种间无关联的0假设。采用统计量W=VRN来检验VR值偏移1的显著程度,如种间不显著关联,则W落入由下面分布给出的界限内的概率有 90%。
2.2.2 成对物种间联结性检验 根据2×2列联表的统计量确定被测成对种间的联结性,公式为:

在下列2种情况下,x2值被认为是有偏差的:(1)2×2列联表中任一小格期望值小于1,(2)多于两小格的期望值小于5。这种偏差可以应用Yates的连续校正系数来纠正[7]。Yates系数纠正公式为:

其中:n为取样总数,a为2个物种均出现的样方数,b和c分别为仅有1个物种出现的样方数,d为2个物种均未出现的样方数。当ad>bc时为正联结,ad>bc时为负联结。若>3.841(0.01<P<0.05),则表示种对间联结性显著;若>6.635(P<0.01)则表示种对间联结性极显著。
2.2.3 种间关联度的测定 本研究采用测定2个物种关联度较好的Ochiai指数,该指数在无关联时等于 0,在最大关联时为 1[8]。Ochiai指数公式为:

3 研究结果
3.1 不同恢复阶段群落主林层的总体关联性分析
群落主林层树种间的整体关联测定结果见表1。表1表明:乔灌过渡阶段、乔林阶段和顶极阶段的群落的种间的总体关联性的方差均值均大于1,但检验统计量W差异较大,经检验分别为无关联性、正关联性和显著正关联性。这一变化规律反映出随退化喀斯特森林自然恢复,树种竞争分化的过程。乔灌过渡阶段,群落垂直结构正处于分化阶段,乔木层不完整,乔木树种和个体少,个体间竞争弱,乔木层种间总体关联性为无关联;乔林阶段,乔木树种多,个体多且密集,同处相同层次,竞争激烈,总体关联性表现为正关联;顶极阶段的群落,乔木层树种经充分竞争分化,个体间形成相互依存的良好关系,总体种间关联性表现为显著正关联性。乔木优势树种种间关联性这一变化,印证了 “随着植被群落演替的进展,群落结构及其种类组成将逐渐趋于完善和稳定,种间关系也将同步趋于正相关,以求得多物种间的稳定共存”[9]的观点。
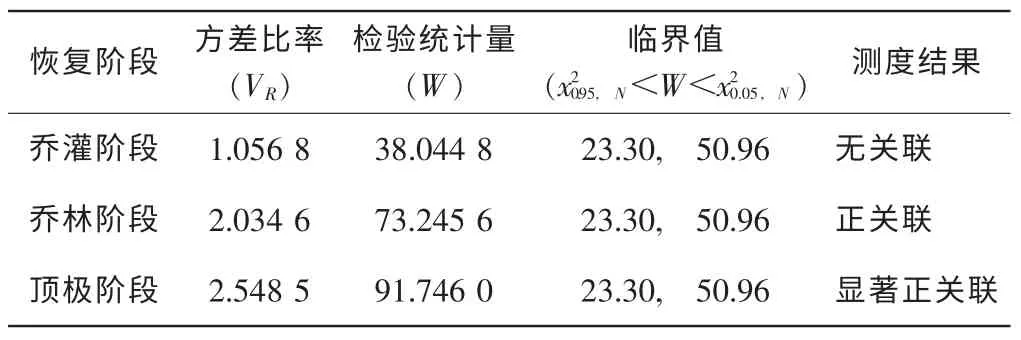
表1 不同恢复阶段群落主林层整体关联性Table1 Overall association of main layers in different recovery stage communities
3.2 不同恢复阶段群落主林层中主要种对间的关联性分析
深入分析群落主林层中主要种对间的关联性是理解群落主林层的整体关联性的基础。对乔灌过渡阶段、乔林阶段和顶极阶段的群落主林层中重要值排名前10位的优势种(表2)种对间的关联性进行测定,结果见表3~6。
正联结反映种间相互依赖,负联结则相互排斥。3个阶段群落主林层主要树种间共135个种对,其中,正联结种对66对,负联结种对60对,联结为0的种对9对,整体表现为种间相互依赖多于排斥,但按恢复阶段分析,各阶段呈现不同的变化。乔灌过渡阶段群落主林层主要树种共45个种对(表3),其中正联结16对,负联结29对,但均未达到显著相关,其联结指数(Ochiai指数)主要集中于0.4以下,种对高达31对(表3),且多为负联结,反映树种随机侵入,种间相互排斥,物种间的分布具有相对独立性的特点。乔林阶段(表4)正联结 19对,负联结26对,种间正相关种对增加,负相关减少,到顶极阶段(表5),正联结31对,负联结14对,正相关最多,占69%,2个阶段的种间联结指数大于0.4的种对分别为19对和22对(表6),还出现呈紧密联系有极显著正联结种对(多脉青冈和长叶木兰,I=0.667)和显著正联结(短萼海桐和天仙果,I=0.820)种对,反映出种间趋向于相互有利、联系紧密的方向发展。因此,这种从负联结低关联性的种对为主,发展到以正联结高关联性种对为主的种对变化规律,反映了从乔灌过渡阶段、乔林阶段和顶极阶段群落主林层树种间关系从相对独立、相互排斥的关系,经种间充分竞争,逐渐形成相互依存关系的发展过程。

表2 不同恢复阶段主要树种重要值Table 2 Important values of main tree species during different recovery stage communities
4 结论与讨论
随着退化喀斯特森林从乔灌过渡阶段经乔林阶段到顶极阶段恢复,主林层种间整体关联呈无关联性→正关联性→显著正关联性,反映了树种间竞争分化的过程,也反映出群落环境由乔灌阶段的不稳定到乔林阶段的不典型,发展至顶极状态群落环境的稳定与典型,也揭示了茂兰喀斯特森林恢复是朝着成熟度高,结构和功能完善的状态发展。
随着退化喀斯特森林从乔灌过渡阶段经乔林阶段到顶极阶段恢复,群落主林层的树种间联结关系从相对独立、相互排斥的关系,经种间充分竞争,逐渐形成相互依存关系的发展过程。
种间的联结性通常被认为是下列原因引起:①相似(正联结)与不相似(负联结)的环境需要;②一个种为另一种创造了定居条件或者前者对后者施加了压力造成了正联结;③两者在竞争资源中互相排斥造成负联结;④两者在根系中通过物理的或化学的因素互相影响也可造成正联结或负联结[10-11]。本研究对茂兰喀斯特森林群落种间联结性研究印证了上述部分关于导致联结性的原因,如顶极阶段的短萼海桐和天仙果呈显著正相关,印证了原因①,因为2个种同处于主林层较低层次,受林冠上层遮蔽,其生长环境相似、稳定且互为补充、相互依赖,为显著正联结,且联结紧密(I=0.820);又如多脉青冈和长叶木兰呈极显著正相关则是原因②的例证,多脉青冈为主林层上层优势种,幼苗耐荫蔽,大树喜光,而长叶木兰处于主林层较低层次,它们对荫蔽度要求依赖于多脉青冈等上层乔木生长造成的荫蔽环境,因而呈极显著的正关联,且联结较紧密(I=0.667)。但是,喀斯特地区因为缺土少水,岩石裸露,树木生长于石缝、石沟、石槽、土面、石面和石洞等小生境上,树种在空间上的分布不仅要受制于小生境的分布影响,而且受制于因缺土少水带来的极端环境的制约,这种极为复杂生境,使得地面环境资源的配置与常态地区相去甚远,而已有的种间联结性认识绝大部分来自于对常态地貌条件下群落种间关系的研究,显然不能满足对喀斯特地区森林群落种间联结的解释,如本研究中3个恢复阶段的135个种对中仅3个种对关联性为显著相关,明显的低于其他常态地貌上所研究的[12-14],难以解释。因此,我们认为,还应从影响树种分布的环境高度异质性、极端环境脆弱性以及树种的生物学、生态学特征以及种间联结性测度方法等方面深入研究,进一步认识群落种间关系。

表3 乔灌过渡阶段群落主林层树种间Ochiai指数和χ2-检验统计量Table 3 Interspecific association Ochiai index and χ2 values of main layer in transitional stage from shrub to tree stage

续表3

表4 乔林阶段群落主林层树种间Ochiai指数和χ2-检验统计量Table 4 Interspecific association Ochiai index and χ2 values of main layer in tree stage

表5 顶极阶段群落主林层树种间Ochiai指数和χ2-检验统计量Table 5 Interspecific association Ochiai index and χ2 values of main layer in climax stage

表6 不同恢复阶段群落主林层种间Ochiai指数(I)分布Table 6 Distribution of Ochiai index(I)of main layers in different recovery stage communities
致谢:研究工作得到了茂兰国家级自然保护区管理局冉景丞、魏鲁明、余登利等和贵州大学林学院安明态、杨瑞、潘柳廷、刘攀峰、龙俊、杨华斌、熊华等的大力支持。特致谢意!
[1]喻理飞.退化喀斯特森林自然恢复的生态学过程研究[D].南京:南京林业大学,1998:17-18.YU Lifei.A Study on Ecological Process of Degraded Karst Forest Natural Restoration[D].Nanjing:Nanjing Forestry U-niversity,1998:17-18.
[2]李援越,祝小科.喀斯特山地次生灌丛优势种群种间联结性的初步研究[M]//朱守谦.喀斯特森林生态研究(Ⅱ)贵阳:贵州科技出版社,1997:136-140.
[3]MULLER D,ELLENBERG H.植被生态学的目的和方法[M].鲍显诚,译.北京:科学出版社,1986:247-252.
[4]孙儒泳,李庆芬,牛翠娟,等.基础生态学[M].北京:高等教育出版社,2002:142-143.
[5]张金屯.数量生态学[M].北京:科学出版社,2004:98-102.
[6]SCHLUTER D A.Variance test for detecting species association with some example applications[J].Ecology,1984,65(3):998-1000.
[7]WANG Bosun,PENG Shaolin.Studies on the measuring techniques of interspecific association of the lower subtropical evergreen-broadleaved forest The exploration and the revision on the measuring formulas of interspecific association[J].Acta Phytoecolt Geobot Sin,1985,9(4):274-288.
[8]LUDWIG J A,REYNOLD J F.Statistical Ecology Primer on Methods and Computing[M].New York:Wiley-interscience Publication,1988:129-131.
[9]DU Daolin,LIU Yuncheng,LI Rui.Studies on the interspecific association of dominant species in subtropical Castanopsis fargesii forest of Jinyun Mountain,China[J].Acta Phytoecol Sin,1995,19(2):149-157.
[10]SMITH G.Quantitative Plaint Ecology[M].3rd.Oxford:BLackwell Scientific Publications,1983:106-128.
[11]KERSHAW K A,LOONEY J H.Quantitative and Dynamic Plant Ecology[M].3rd.London:Edward Arnold Limited,1985:78-94.
[12]哀建国,翁国杭,董蔚.石垟森林公园常绿阔叶林主要种群的种间联结性[J].浙江林学院学报,2008,25(3):324-330.AI Jianguo,WEN Guohang,DONG Wei.Interspecific association of primary plant populations in an evergreen broadleaf forest at Shiyang Forest Park of Zhejiang Province[J].J Zhejiang For Coll,2008,25(3):324-330.
[13]李登武,刘国彬,张文辉,等.秦巴山地栓皮栎所在群落主要乔木树种种间联结性的研究[J].西北植物学报,2003,23(6):901-905.LI Dengwu,LIU Guobin,ZHANG Wenhui,et al.A study on interspecific association of principal tree species in the communities including Quercus variabilis on the Qinling and Bashan Mt.[J].Acta Bot Boreal-Occident Sin,2003,23(6):901-905.
[14]邓贤兰,刘玉成.井冈山自然保护区栲属群落优势种群的种间联结关系研究[J].植物生态学报,2003,27(4):531-536.DENG Xianlan,LIU Yucheng.Interconnection among dominate plant populations of Castanopsis community in Jinggang Mountain Nature Reserve[J].Acta Phytoecol Sin,2003,27(4):531-536.
