远缘植物试管嫁接及ISSR分析
2010-12-05邵果园陆方方
邵果园,陆方方
(浙江农林大学 农业与食品科学学院,浙江 临安 311300;2.西南大学 园艺园林学院,重庆 400715)
试管嫁接又称微嫁接,是在试管内将砧木与接穗进行嫁接获得新植株的技术[1-2],是以组织培养为基础,嫁接植物在人工控制的环境条件下培养,排除了外界环境因素对嫁接成活的影响,已被广泛地应用于许多植物的研究与生产中。远缘嫁接是不同属不同种甚至是不同科的植物间进行嫁接,如嫁接成功,不仅能够提高观赏价值,提高产量和改进品质,还能培育出新品种[3-4]。由于远缘嫁接砧穗的不亲和性等因素减小了远缘嫁接成功的可能性,现有的研究还仅限于草本与草本植物、木本与木本植物之间的嫁接,而且对嫁接变异现象还没有充分的遗传理论基础[5-6],为此,本研究采用试管嫁接的方法,将大岩桐Sinningia speciosa,鸡冠花Celosia cristata,长寿花Kalanchoe blossfeldiana,黄色马蹄莲Zantedeschia elliottiana,百合Lilium longiflorum,苹果Malus domestica和阳桃Averrhoa carambola等7种植物进行组合嫁接,以期对双子叶与单子叶植物、草本与木本植物远缘嫁接模式进行新的尝试,并利用简单重复序列区间扩增多态性(inter-simple sequence repeat,ISSR)分子标记技术,对成活的试管嫁接苗进行分子水平的检测,以期获得嫁接变异在DNA方面的证据,从而增加嫁接引起可遗传的变异的理论支持,也为后续的研究提供实验技术参考。
1 材料与方法
1.1 试验材料
试材有大岩桐(苦苣苔科Gesneriaceae苦苣苔属Sinningia),鸡冠花(苋科Amaranthaceae青葙属Celosia),长寿花(景天科Crassulaceae伽蓝菜属Kalanchoe),黄色马蹄莲(天南星科Araceae马蹄莲属Zantedeschia),百合(百合科Liliaceae百合属Lilium),苹果(蔷薇科Rosaceae苹果属 Malus)及阳桃(酢浆草科Oxalidaceae阳桃属Averrhoa),均为组织培养无菌体系,由西南大学园艺园林学院果树学重点实验室建立。
1.2 试验方法
1.2.1 嫁接 试验设计18对嫁接组合。在超净工作台上,用磨尖的镊子在砧木上面戳一个深3~5 mm的孔,然后把下端表皮刮出伤口的接穗插入砧木上的孔中。嫁接苗接种于适合砧木增殖的培养基。接种30株·组合-1,重复3次。每天观察砧木接穗的生长状态及结合部情况,40 d后统计成活率及生长情况。
1.2.2 ISSR分析 ①基因组DNA的提取:取嫁接成活植株及相应接穗、砧木的嫩叶1~2片,约0.5 g,采用王飞等的方法[7]提取基因组 DNA,用 8 g·kg-1琼脂糖凝胶电泳检测,终质量浓度调整至10 mg·L-1。②ISSR扩增:本研究采用多重优化ISSR反应体系,20 μL反应体系中各组分的浓度分别为:1×聚合酶链式反应(PCR)缓冲液,0.125 mmol·L-1三磷酸碱基脱氧核苷酸(dNTP),2 mmol·L-1氯化镁,1×16.67 nkat Taq DNA聚合酶,0.3 μmol·L-1引物和 20 ng模板。PCR反应程序为 94℃预变性 4 min,然后94℃变性1 min,50℃退火45 s,72℃延伸1.5 min,35个循环,72℃后延伸10 min。扩增产物用15 g·kg-1琼脂糖凝胶电泳检测。随后进行条带统计及分析。
2 结果与分析
2.1 植物远缘嫁接的成活率
试验结果表明(表1),远缘植物之间的不同嫁接组合,嫁接的成活率有很大的差异。草本植物之间的所有嫁接组合均有不同程度的成活率,且差异显著,最高成活率可达33.3%;单子叶与双子叶植物的组合中,黄色马蹄莲(接穗)+长寿花(砧木)、百合 +大岩桐组合没有成活植株,百合 +长寿花嫁接成活率较高,达15.0%;草本与木本植物的嫁接组合均未有成活植株。造成差异的主要原因可能是由于不同植物间亲缘关系的远近造成的嫁接亲和性的不同[8],同时,与嫁接方法和嫁接技术的娴熟度也有一定关系。
2.2 远缘嫁接苗的生长状况
成活的远缘植物嫁接苗普遍表现出生长缓慢、衰弱或畸形。嫁接后45 d,鸡冠花 +大岩桐的接穗在瓶内已经开花,但生长逐渐衰弱(图1-1),从其接穗鸡冠花上取茎段进行增殖,增殖后代的叶片边缘及叶脉出现细微的红线(图1-2),这与原株略有不同,且株型呈莲座状(图1-3),伸长生长极缓慢。
2.3 远缘植物嫁接的ISSR分析
从100条ISSR引物中筛选出在接穗和砧木中有差异,且多态性明显的引物用于检测分析。嫁接苗ISSR检测结果统计见表2,可见嫁接苗扩增位点存在以下3种情况(图2):①偏亲性位点。按照传统遗传学的观点,嫁接后代基因组应该和接穗基因组一致,能够保持母本的特性。长寿花+大岩桐、长寿花 +鸡冠花2个组合在嫁接后,接穗所扩增出的位点和接穗来源株一致,未发生变化。②非亲性扩增位点。大岩桐 +鸡冠花、大岩桐 +长寿花、鸡冠花 +大岩桐、大岩桐 +鸡冠花等4个组合嫁接后一些接穗的扩增条带与接穗和砧木来源株相比较,表现位点缺失或者出现新的位点。这种变化可能是由于远缘嫁接所致。③双亲杂合位点。嫁接苗杂合接穗与砧木来源株的位点,如鸡冠花 +大岩桐、大岩桐+鸡冠花嫁接成活苗。这说明,嫁接后砧木与接穗产生相互影响,可能发生了大分子物质的交流,从而使砧木的某些遗传信息整合到了接穗的遗传物质中。

表1 植物嫁接组合结果统计Table 1 Result of grafting combinations

图1 鸡冠花+大岩桐嫁接苗的几种异常生长状况Figure 1 Unusual growth of the grafting plants from Celosia cristata(scion)+ Sinningia speciosa(rootstock)
产生位点变化的嫁接苗,初始外观性状表现与原来差异并不大。对鸡冠花+大岩桐组合的接穗进行增殖,并对增殖后代进行ISSR检测,发现来自砧木的条带在增殖后代中仍然存在。

表2 嫁接苗ISSR检测结果Table 2 Result of ISSR analysis of grafting plants
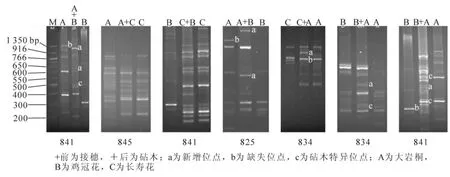
图2 不同嫁接苗在7个引物下的扩增条带Figure 2 Result of ISSR amplification
3 讨论
3.1 远缘试管嫁接亲和力与成活率的关系
亲和力被认为是嫁接成活的基本条件,植物嫁接的亲和性机制问题也是目前嫁接理论研究中的热点。接穗与砧木在分类学上的关系愈近,嫁接成功率就越高;分类学上的关系愈远,排斥现象愈强烈,嫁接就越不容易成功。在本研究中,不同嫁接组合的嫁接成活率各不相同,差异也比较大,从草本与木本植物、单子叶与双子叶植物、草本植物之间嫁接成活率来看,草本与木本植物之间由于其组织结构,内部代谢等方面都大相径庭,亲和性比较差,因而嫁接成功的难度非常大。相对而言,草本植物之间的嫁接成活率要大一些,单子叶和双子叶植物间也有少量成活的嫁接苗。另外,远缘试管嫁接的成活率还与嫁接组合,嫁接技术条件,嫁接方法,砧穗形态大小,甚至试验操作者的熟练程度等多种因素相关[8],因而它并不能够作为亲和力强弱的绝对指标。
3.2 嫁接变异的原因
嫁接可以引起可遗传的变异,但嫁接变异的机制至今还不确定,过去在有关远缘嫁接遗传变异的研究中,都推测可能是砧木中的某些DNA片段被转运到接穗中所致[9-10],但至今未获得此方面直接的分子生物学证据[11]。McClintock[12]最早提出了转座子的概念,并进行了系统的研究,正常情况下,转座子不表现活性或活性很低,但创伤、原生质体分离、组织培养和病毒、细菌或真菌的感染等环境压力常可激活转座子转座,使基因发生重排,适应逆境的突变存活下来。本研究中,用ISSR分析检测到几种嫁接组合均在多种引物下出现了有差异的位点,鸡冠花+大岩桐与大岩桐+鸡冠花组合嫁接后,接穗及增殖后代甚至出现了砧木的特异带纹,这就支持了传统的砧木与接穗之间有可能是发生了DNA等大分子物质的转移,产生了基因交流的观点。但试验结果所示的位点新增与缺失等现象,或许也与远缘嫁接逆境引起转座子活动而导致基因突变有关。
[1]JONARD R,HUGARD J,MACHEIX J J,et al.In vitro micrografting and its applications to fruit science[J].Sci Hortic,1983,20(1):147-159.
[2]ESTRADA-LUNA A A,LóEZ-PERALTA C,CáDENAS-SORIANO E.In vitro micrografting and the histology of graft union formation of selected species of prickly pear cactus(Opuntia spp.)[J].Sci Hortic,2002,92(3):317-327.
[3]李保印,刘用生,周秀梅,等.柳属与杨属植物远缘嫁接研究[J].生物学通报,2004,39(10):19-20.LI Baoyin,LIU Yongsheng,ZHOU Xiumei,et al.The preliminary research on the distant grafting of Salix and Populus[J].Bull Biol,2004,39(10):19-20.
[4]孟昭璜.远缘嫁接无性杂交培育小麦新品种的方法:中国,CN1765175[P].2006-05-03.
[5]刘用生.对 “接穗郁李影响杏砧变异的实验”的解释[J].遗传,2000,22(6):401-402.LIU Yongsheng.The explanation for phenotype variation by the action of scion Prunus japonica Thunb on stock Prunus armeniaca L.[J].Hereditas,2000,22(6):401-402.
[6]夏涵涵,杨敏,何波,等.远缘植物试管苗嫁接及ISSR分析[J].西南农业学报,2007,20(6):1180-1183.XIA Hanhan,YANG Min,HE Bo,et al.Study on distant grafting in vitro and ISSR markers[J].Southwest China J Agric Sci,2007,20(6):1180-1183.
[7]王飞,巩振辉,马维,等.一种适宜PCR检测的番茄DNA微量提取方法[J].西北农业学报,2006,15(1):169-172.WANG Fei,GONG Zhenhui,MA Wei,et al.A new method of mini-preparation DNA for tomato suited to PCR fetection[J].Acta Agric Boreali-Occident Sin,2006,15(1):169-172.
[8]刘用生,李保印,李桂荣,等.嫁接杂交与果树遗传的特殊性[J].遗传,2004,26(5):705-710.LIU Yongsheng,LI Baoyin,LI Guirong,et al.Graft hybridization and the specificity of heredity in fruit trees[J].Hereditas,2004,26(5):705-710.
[9]TALLER J,HIRATA Y,YAGISHITA N,et al.Graft-induced genetic changes and the inheritance of several characteristics in pepper(Capsicum annuum L.)[J].Theor Appl Genet,1998,9(7):705-713.
[10]齐明.运用ISSR分子标记鉴定杉木×侧柏远交杂种[J].浙江林学院学报,2008,25(5):666-669.QI Ming.Hybrid identification of Cunninghamia lanceolata × Platycladus orientalis based on ISSR markers[J].J Zhejiang For Coll,2008,25(5):666-669.
[11]ZHANG Danhua,MENG Zhaohuang,XIAO Weiming,et al.Graft-induced inheritable variation in mungbean and its application in mungbean breeding[J].Acta Bot Sin,2002,44(7):832-837.
[12]崔克明.植物发育生物学[M].北京:北京大学出版社,2007:250-251.
