骨髓间充质干细胞条件培养液对小鼠卵母细胞的孤雌激活作用
2010-09-11冯定庆周颖李彩荣高婷凌斌
冯定庆,周颖,李彩荣,高婷,凌斌,
(1.安徽医科大学附属省立医院分子医学重点实验室,合肥 230001; 2.安徽医科大学附属省立医院妇产科,合肥 230001)
研究报告
骨髓间充质干细胞条件培养液对小鼠卵母细胞的孤雌激活作用
冯定庆1,周颖1,李彩荣1,高婷2,凌斌1,2
(1.安徽医科大学附属省立医院分子医学重点实验室,合肥 230001; 2.安徽医科大学附属省立医院妇产科,合肥 230001)
目的研究骨髓间充质干细胞(mesenchymal stem cell,MSC)条件培养液对小鼠M II卵母细胞的孤雌激活作用及胚胎发育能力。方法分离、培养小鼠MSC,获取MSC条件培养液(conditioned medium of MSC,CM)。通过促排技术获取小鼠MII卵母细胞,分别采用CM、7%乙醇、IVF方法激活,体视显微镜下观察原核形成及囊胚形成率。在CM激活后不同时间点,利用α/β-tubulin抗体标记纺锤体,激光共聚焦显微镜下观察有/无细胞松弛素B(CB)存在时纺锤体的运动变化。结果CM可以激活小鼠MII卵母细胞,最佳刺激时间为40 min,激活率达到95.4%,囊胚形成率为62%,与7%乙醇组比较无显著差异,但明显低于IVF组(95.4%vs.100%;62%vs. 88%,P<0.01)。CB可以抑制纺锤体的旋转,阻止第二极体的排出,促进二倍体孤雌胚形成,提高囊胚形成率(62%vs.9%,P<0.01)。结论CM能有效激活小鼠M II卵母细胞并促进孤雌发育。
间充质干细胞;卵母细胞;孤雌激活;细胞松弛素B

注:A. CM处理40 min后;B.CM激活后CB处理2 h;C. CM激活后CB处理4 h;D.异常纺锤体图1CM激活卵母细胞的纺锤体运动(标尺= 20 μm)Note: A. 40 min after CM treatment; B. 2 h after CB treatment following activation; C. 4 h after CB treatment following activation; D. abnormal mitotic sPindle comPleXeFig.1Dynamic changes of the sPindle comPleXes of CM activated oocytes (Scale bar = 20 μm)
卵母细胞的孤雌激活是核移植的关键技术,也是研究哺乳动物孤雌生殖及受精机制的重要手段之一。引起卵母细胞激活的因素很多,通常可分为物理刺激和化学刺激两类。前者有电刺激和温度刺激等[1];后者常用的试剂有乙醇、离子霉素、放线菌酮(CHX)、二甲基氨基嘌呤(DMAP)、Ca2+载体等[2-4]。不同的激活方法和激活剂的选择,对卵母细胞的孤雌激活率、卵裂率及胚胎发育均有较大程度的影响。
骨髓间充质干细胞(mesenchymal stem cells,MSC)是一种存在于骨髓内的成体干细胞,除了具有自我更新和多向分化潜能外,还可以分泌EGF、LIF、TGF-β、FGF-2、VEGF、MCP-1等多种细胞因子,发挥营养作用,参与构成造血微环境[5,6]。我们的前期研究发现,MSC条件培养液(CM)能够促进小鼠体外培养的卵泡发育,卵母细胞核浆同步成熟,提高成熟率和卵母细胞质量[6]。进一步研究显示,成熟卵母细胞长时间暴露于CM中有激活及卵裂现象的发生。为了探讨最佳激活条件及激活效率,本实验将小鼠MII卵母细胞用CM处理不同时间,观察激活效率及孤雌胚胎发育情况,从而对CM孤雌激活作用做出评价。
1 材料与方法
1.1 实验动物
清洁级昆明种小鼠,6~8周龄,体质量18~22 g,来源于安徽医科大学实验动物中心【SCXK(皖) 2005-001】,并按动物使用的3R原则给予人道的关怀。其中,雌鼠82只用于促排获取MII卵母细胞和MSC,雄鼠6只用于获取精子进行体外受精。动物自由摄食,维持8:00~20:00光照循环,室温20~25℃,湿度40%~60%。
1.2 主要试剂与仪器
DMEM、胎牛血清(FCS,Gibco,美国),HTF (LifeGlobal,美国),M16、透明质酸酶、细胞松弛素B (CB),α/β-tubulin抗体均购自美国Sigma公司,蛋白酶E(BioShare,德国),Percoll(Amersham Bioscience,瑞典),FITC-anti-CD11b、FITC-anti-CD34、PE-cy5-anti-CD44、PE-anti-CD105(eBioscience,美国),HCG、PMSG(杭州动物药品厂,中国),4孔培养板(Nunc,Roskilde,丹麦),流式细胞仪(FACS,美国Beckman),SZX7体视显微镜、恒温载物台(Olympus,日本),激光共聚焦显微镜(LSM510,德国Zeiss)。
1.3 M SC分离、鉴定及条件培养液的获取
颈椎脱臼处死小鼠,无菌条件下取出股骨和肱骨,PBS反复冲洗骨髓腔;收集冲洗液缓慢加到60%Percoll液面上,2000 r/m in离心20 m in;收集单个核细胞,PBS洗涤2次后用含10%FCS的DMEM重悬,5×106细胞/m L密度接种培养瓶,3 d后换液弃去未贴壁细胞,获得原代MSC,CD11b、CD34、CD44、CD105标记细胞,FACS鉴定。F1代MSC 90%铺满瓶底时换液,培养48 h后收集上清即为CM,0.22 μm滤膜过滤即用。
1.4 M II卵母细胞获取及激活
雌鼠腹腔注射PMSG 7.5 IU,48 h后注射HCG 7.5 IU;14 h后处死小鼠,分离双侧输卵管放入37℃预温的M16培养基内,体视显微镜下针头刺破输卵管壶腹部,获得卵丘-卵母细胞复合体(COCs); 0.1%透明质酸酶消化去除颗粒细胞,HTF洗涤后用于激活处理。
卵母细胞的激活分别采用三种方法:①CM,MII卵母细胞用新鲜收获的CM处理0 m in、10 m in、40 m in、1 h、2 h;含10%FCS DMEM处理的MII卵母细胞设为试剂对照;②7%乙醇,MII卵母细胞用含7%乙醇的CZB培养基刺激6 min;③体外受精(IVF),从雄性小鼠获取精子,获能1~1.5 h后,调整精子密度为0.7×106~1.3×106精子/m L,与用0.5%蛋白酶E去除透明带的MII卵母细胞进行体外受精。
CM和乙醇激活的卵母细胞用7.5 μg/m L CB处理4 h,然后所有激活的卵母细胞用含0.4%BSA的CZB培养基培养96 h,观察胚胎发育情况。其中,48 h时向培养基内添加终浓度为1 mmol/L的谷氨酰胺和5.56 mmol/L的葡萄糖。
1.5 激光共聚焦显微镜观察纺锤体运动
收获激活后0、1、2、4、6、12、16、20、24 h的卵母细胞,3.7%多聚甲醛固定30 min放入封闭液(含1%BSA、2%山羊血清、0.1 mol/L甘氨酸、0.01% Triton X-100、0.2%叠氮钠)内4℃保存;α/β-tubulin抗体37℃孵育1 h,FITC标记二抗作用45 min,然后10 μg/m L PI孵育10 min,其间每步均用0.1% PVP/PBS洗涤;最后70%甘油封片,激光共聚焦显微镜观察结果。
1.6 统计学处理
2 结果
2.1 M SC的形态特征及表型
密度梯度离心结合贴壁筛选法可以获得纯度很高的MSC,原代培养3d换液可见贴壁细胞呈梭形、纺锤形,增殖迅速,传代后形态更为均一。FACS检测,MSC表型为:CD11b-,CD34-,CD44+,CD105+,与MSC公认表型特征一致(参见前期结果,文献6)。
2.2 CM对M II卵母细胞的激活作用
CM处理10 min后46.5%的卵母细胞被激活,而处理40 m in、1 h、2 h激活率均为95%左右,但是随着时间延长,胚胎碎片的数目显著增加。因此,CM最佳作用时间为40 m in。与乙醇组比较激活效率无明显差异;两组与IVF组比较差异有统计学意义,P<0.01。试剂对照组有6.3%的卵母细胞自发激活,MII卵母细胞自发激活常见于低等脊椎动物和两栖动物[7],因此后续实验未予以分析。见表1。
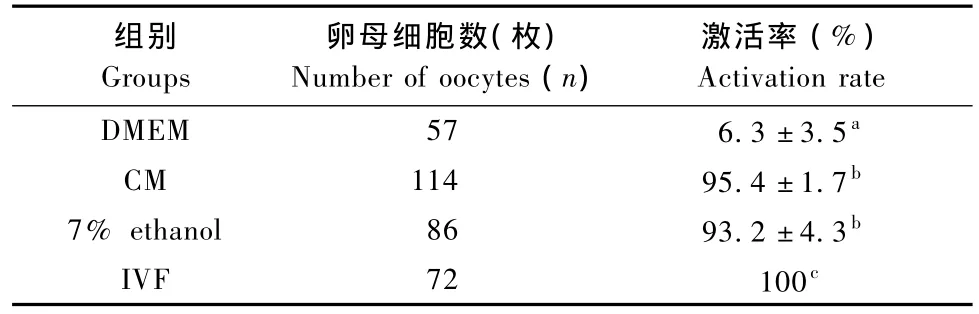
表1 不同方法对小鼠M II卵母细胞的激活效率比较Tab.1 Activation rate of mouse MII oocytes treated by different protocols
2.3 不同激活方法对胚胎发育的影响
CM激活的卵母细胞经CB处理后,80%形成2个原核,未加CB组大多数形成1个原核或快速发生卵裂;CB对激活率无明显影响(97.2%vs. 95.4%),但囊胚形成率差异显著(62%vs.9%,P<0.01)。CM、7%乙醇激活的MII卵母细胞经CB处理后,囊胚形成率均为62%左右,显著低于IVF组(62%vs.88%,P<0.01)。见表2。

表2 不同激活方法对小鼠胚胎发育的影响Tab.2 The effect of different treatments on the development of mouse embryos
2.4 CM活化的卵母细胞纺锤体的运动变化
MII卵母细胞受CM作用40 min后恢复减数分裂,纺锤体平行于细胞膜,染色质排列在赤道板上(图1A);未加CB处理,纺锤体发生旋转,活化1~2 h后与细胞膜垂直,染色质被拉到两极,随后排出第二极体;经过CB处理,减数分裂的恢复和姊妹染色体分离不受影响,但纺锤体运动被抑制,不能排出第二极体(图1B);CB作用4 h后形成2个原核(图1C)。但是CM活化的卵母细胞无论是否经CB处理,8%左右1-细胞胚胎形成异常纺锤体,如3极或4极纺锤体、中心小体异位、染色质排列紊乱等(图1D)。图1见彩插4。
3 讨论
哺乳动物卵母细胞的激活是生殖过程、IVF、ISCI及核移植技术成功实施的关键因素之一,其中卵母细胞的孤雌激活更是为克隆研究、父源与母源基因组在胚胎发育中的作用、印记基因的表达及胚胎干细胞的获取创造了条件。目前,已经建立了多种物理的、化学的人工激活方法[1-4],但是关于间充质干细胞CM液对卵母细胞的激活作用尚未见报道。
MSC通过旁分泌/自分泌作用分泌一系列细胞因子,在局部发挥重要的生物学作用,包括抑制局部免疫反应、阻止纤维化形成、抑制细胞凋亡、促进血管形成、刺激组织细胞/干细胞有丝分裂和分化参与内源性修复[5]。大量的研究发现,其中的一些因子能够直接促进卵母细胞发育成熟,提高胚胎发育能力,有些就存在于卵泡液中由卵母细胞和颗粒细胞共同分泌[8,9];我们前期的研究结果也证实了这一点[6]。LIF有促进很多动物孤雌胚/IVF胚胎原核形成及分裂发育、提高内细胞群数量等作用[10-12]。在卵泡发育的各个阶段,卵母细胞和颗粒细胞持续表达EGF受体;O’Donnell等[13]报道,EGF、TGF-α与EGF受体结合使受体胞内部分发生磷酸化,可以提高卵母细胞胞质[Ca2+]i水平,增加细胞通透性,这是卵母细胞活化最重要的早期事件[14]。我们的结果显示,CM可以有效激活小鼠MII卵母细胞,最佳刺激时间为40 m in,激活效率达到95%左右;随着刺激时间延长,激活效率不再增加,而胚胎内碎片的数量却明显增加;囊胚形成率为62%,与7%乙醇组比较均无显著差异。MII卵母细胞用含10%FCS的DMEM处理,也有6.3%左右发生活化,但是均不能发育到囊胚阶段,我们认为是自发性活化,这在低等脊椎动物和两栖动物较为常见,并且老化的卵母细胞更易发生[7]。
CB是一种促进微丝解聚药物,可以抑制胞质分裂和纺锤体的旋转运动,阻止第二极体的排出,但不影响姊妹染色体的分离,被广泛用于克隆、获取二倍体或多倍体胚胎[15]。CM激活MII卵母细胞后,用7.5 μg/m L CB处理4 h,所形成的基本上为二倍体孤雌胚,否则多为单倍体;后者卵裂速度明显快于二倍体孤雌胚,但是囊胚形成率显著低下(9%vs. 62%)。在CM激活组,不论CB处理与否,约有8%左右1-细胞胚胎出现异常纺锤体,有的还形成假纺锤体结构,具体原因还不清楚。
上述实验结果显示,CM能够有效激活MII卵母细胞,即恢复减数分裂、原核形成并发育至囊胚。但是CM成分复杂,阐明具体的作用机制及其在核移植、获取无伦理争议的胚胎干细胞等领域的应用,还需要进一步的研究。
(本文图1见彩插4。)
[1]Fissore RA,Robl JM.Intracellular Ca2+response of rabbit oocytes to electrical stimulation[J].Mol Reprod Dev,1992,32:9-16.
[2]Yang X,Presicce GA,Moraghan L,et al.Synergistic effect of ethanol and cycloheximide on activation of freshly matured bovine oocytes[J].Theriogenology,1994,41:395-403.
[3]Ma SF,Liu XY,Miao DQ,et al.Parthenogenetic activation of mouse oocytes by strontium chloride:a search for the best conditions[J].Theriogenology,2005,64:1142-1157.
[4]Yi YJ,Park CS.Parthenogenetic development of porcine oocytes treated by ethanol,cycloheximide,cytochalasin B and 6-dimethylaminopurine[J].Anim Reprod Sci,2005,86:297-304.
[5]Cap lan AI,Dennis JE.Mesenchymal stem cells as trophic mediators[J].J Cell Biochem,2006,98:1076-1084.
[6]Ling B,Feng DQ,Zhou Y,et al.Effect of conditioned medium of mesenchymal stem cells on the in vitro maturation and subsequent development of mouse oocyte[J].Braz J Med Biol Res,2008,41:978-985.
[7]Krivokharchenko A,Popova E,Zaitseva I,et al.Development of parthenogenetic rat embryos[J].Biol Reprod,2003,68:829-836.
[8]Sutton ML,Gilchrist RB,Thompson JG.Effects of in-vivo and in-vitro environments on the metabolism of the cumulus-oocyte complex and its influence on oocyte developmental capacity[J]. Hum Reprod Update,2003,9:35-48.
[9]Chian RC,Buckett WM,Tan SL.In-vitro maturation of human oocytes[J].Reprod Biomed Online,2004,8:148-166.
[10]Tsai HD,Chang CC,Hsieh YY,et al.Recombinant human leukemia inhibitory factor enhances the development of preimplantation mouse embryo in vitro[J].Fertil Steril,1999,71:722-725.
[11]Dunglison GF,Barlow DH,Sargent IL.Leukaemia inhibitory factor significantly enhances the blastocyst formation rates of human embryos cultured in serum-free medium[J].Hum Reprod,1996,11:191-196.
[12]Ptak G,Lopes F,Matsukawa K,et al.Leukaemia inhibitory factor enhances sheep fertilization in vitro via an influence on the oocyte[J].Theriogenology,2006,65:1891-1899.
[13]O’Donnell JB Jr,Hill JL,Gross DJ.Epidermal growth factor activates cytosolic[Ca2+]elevations and subsequent membrane permeabilization in mouse cumulus-oocyte complexes[J]. Reproduction,2004,127:207-220.
[14]Tomashov-Matar R,Tchetchik D,Eldar A,et al.Strontiuminduced rat egg activation[J].Reproduction,2005,130:467-474.
[15]Zhu ZY,Chen DY,Li JS,et al.Rotation of meiotic spindle is controlled by microfilaments in mouse oocytes[J].Biol Reprod,2003,68:943-946.
Effects of Conditioned M edium of M esenchym al Stem Cells on M ouse Oocyte Parthenogenetic Activation
FENG Ding-qing1,ZHOU Ying1,LI Cai-rong1,GAO Ting2,LING Bin1,2
(1.Anhui Province Key Laboratory of Molecular Medicine,Anhui Provincial Hospital Affiliated to Anhui Medical University,Hefei 230001,China; 2.Department of Obstetrics and Gynecology,Anhui Provincial Hospital Affiliated to Anhui Medical University,Hefei 230001,China)
ObjectiveTo investigate the effects of conditioned medium of mesenchymal stem cells(CM)on the parthenogenetic activation and development of mouse MII oocytes.MethodsMouse mesenchymal stem cells(MSC)were isolated and further cultured to collect CM.Mouse MII oocytes were collected with superovulation and activated with CM,7%ethanol,or in vitro fertilization(IVF),respectively.Pronuclei formation and embryo development were evaluated under the view of a stereomicroscope.At various time-points after activation with or without the presence of cytochalasin B (CB),the dynamic changes of meiotic spindle,as visualized in preparations stained for α/β-tubulin and chromatin,were observed by fluorescent confocal microscopy.ResultsCM effectively activated MII oocytes with the optimal time of 40 min.The rates of activation and blastocyst showed no significant difference between CM and 7%ethanol but were significantly lower in both compared with that in the IVF group(95.4%vs.100%,P<0.01;62%vs.88%,P<0.01).CB inhibited the spindle rotation and polar body extrusion but did not affect chromosomal movement and nuclear division and,thus,facilitated the diploid formation and blastocyst rate(62%vs.9%,P<0.01).ConlusionCM is an effective medium for mouse MII oocyte activation and subsequent embryo development.
Mesenchymal stem cell;Oocyte;Parthenogenetic activation;Cytochalasin B;Mouse
Q-813.7
A
1005-4847(2010)04-0304-04
2009-09-03
国家自然基金(编号:34071805,30901606);安徽省科技攻关项目(编号:06013124B)。
冯定庆(1977-),男,助理研究员,硕士;研究方向:生殖免疫。E-mail:feng.dingqing@gmail.com
凌斌。E-mail:lingbin.ling@gmail.com
