岷江上游山地森林/干旱河谷交错带退耕还林后土壤养分变化和微生物分布特征*
2010-08-02易海燕宫渊波陈林武唐春香黄正全
易海燕,宫渊波,陈林武,2,张 敏,唐春香,黄正全,陈 鑫
(1.四川农业大学长江上游林业生态工程省级重点实验室,四川雅安625014;2.四川省林业科学研究院,成都610081)
岷江上游干旱河谷区是以干旱河谷为基带的一类较特殊的山地生态系统。在干旱河谷山地垂直带谱中,从上至下表现出明显的高山草甸、高山灌丛、亚高山针叶林、干旱河谷灌丛等生态类型[5],山地森林/干旱河谷交错带是该区由土壤水分垂直变化影响而形成的一种特殊的生态交错区。由于该区河流深切及南北走向的横断山,致使东南太平洋季风和西南印度洋季风两股气流受到阻拦,从南向北水分和热量不断降低,由暖湿气流带来的水分被高大山体阻隔在迎风面形成降雨,在背风面的峡谷中气流下沉绝热增温即“焚风”效应,导致河谷气候呈变干变暖的趋势[1-3]。加之山体坡面土壤侵蚀和河流下切作用强烈,是青藏高原东部生态最脆弱、环境最恶劣的地段之一。岷江上游干旱河谷生态系统的脆弱性决定了干旱河谷具有低阈值生态安全和高风险生态退化的特点,是在同区域山地生态系统中最脆弱,存在问题最多,也是植被恢复最关键和最困难的一种特殊地域类型[4]。近几十年来,由于长期的森林过伐、坡地开垦及过度放牧等,导致山地森林破坏极为严重,同时也使干旱河谷的边界向更高的海拔和上下游发展,干旱河谷的林线上移迅速,部分山体森林植被完全消失,干旱化加剧,植被恢复极为困难。利用山地森林/干旱河谷交错带边缘效应的作用,开展植被恢复的试验研究,是干旱河谷区植被恢复的有效途径之一[4]。
土壤养分含量直接影响植物的生长,土壤微生物通过分解动植物残体而参与森林生态系统的能量流动和物质循环,影响植物的生长发育,是土壤肥力的重要指标之一[6-7]。在岷江上游有关土壤养分及微生物特征的研究多集中在亚高山森林和干旱河谷区[8-14],对山地森林/干旱河谷交错带的类似研究少见报道。本文针对岷江上游干旱河谷与山地森林交错带水土流失严重和生态退化的特点,选择典型山地森林/干旱河谷交错带进行土壤养分与微生物特征的研究,以期了解该地区植被恢复过程中对土壤肥力变化的影响,为岷江上游干旱河谷区的植被恢复提供一些基本依据。
1 材料与方法
1.1 研究区概况
研究区设在四川省西部理县干旱河谷的典型地带杂谷脑河支流的甘堡乡熊耳山,位于31°31′06″-31°32′10″N,103°12′25″-103°13′36″E,是一典型的山地森林/干旱河谷交错带,平均海拔2 700m。年均气温11.0℃,>0℃积温3 800~4 500℃,>10℃的有效积温3 200~3 800℃ ,无霜期190 d,年干燥度1.6~2.5,全年日照时数在1 200~2 000 h,年降水量为400~600 mm,蒸发量为降水量的2~4倍。
1.2 土样的采集
于2008年8月在研究区范围内按3个地段选择样地(见表1)。按不同植被类型,海拔从低到高分别为封禁造林地、白杨-旱柳混交林、灌木林、针阔混交林、次生林。其中白杨-旱柳混交林和针阔混交林为开展退耕还林工程后于2002年营造的人工林,次生林和灌木林则为自然恢复的植被类型。在每个样地内每隔5 m按等边三角形设置3个采样点挖土壤剖面,每个剖面分0-10 cm,10-20 cm分层采集土样,土样混合后冷藏并迅速带回实验室,一份鲜样去杂后贮藏于4℃的冰箱内,供微生物数量的测定;另一份风干、去杂、过筛后供土壤养分的测定。

表1 样地基本情况
1.3 土壤样品的分析
土壤有机碳用重铬酸钾氧化-外加热法测定;pH 用pH原位器测量;全N用凯式定氮法测定;水解氮测定采用碱解扩散法;氢氧化钠碱熔钼锑抗比色法测定全P;双酸浸提法测定有效磷;氢氧化钠碱熔,火焰光度计法测定全钾;醋酸铵浸提,火焰光度法测定速效钾[15]。土壤微生物分析:主要采用稀释平板分析法测定细菌、放线菌、真菌3大类群的数量,细菌用牛肉膏蛋白胨培养基;放线菌用改良高氏一号培养基;真菌用马铃薯-蔗糖琼脂培养基。每克干土中菌数=(菌落平均数×稀释倍数)/干土重(g)。土壤生理群微生物的测定:主要采用最可能数值(MNP)法,固氮菌用改良阿须贝无氮培养基;氨化细菌用蛋白胨培养基;好气性纤维素分解菌采用依姆歇涅茨基纤维素分解菌培养基[16]。
2 结果与分析
2.1 山地森林/干旱河谷交错带土壤养分特征
山地森林/干旱河谷交错带土壤养分见表 2。从表2可以看出土壤pH值为干旱河谷地段>交错带山地森林下缘地段>交错带山地森林段;各土层之间pH相比较,除封禁造林地外,其他植被类型pH呈弱酸性并随土层的加深而增大,封禁造林地由于土壤蒸发量大,导致下层土壤水分沿毛细管上升,可溶性盐基离子随之上升到土壤表层,盐基离子在土壤表层积累,从而导致土壤pH 呈弱碱性,并随土层的加深而减小。随着海拔的升高和植被的恢复,土壤水分蒸发量减小、淋溶作用加强,导致土壤呈弱酸性并随土层的加深而增大。由此也可看出交错带下缘土壤及水分条件优于较低海拔的荒地,恢复森林植被的难度也相对较小。次生林土壤的有机碳、全氮、速效氮、速效磷含量最高;全磷含量以灌木林土壤最高;全钾、速效钾的含量以白杨-旱柳混交林土壤最高;封禁造林地的土壤养分含量均处于最低水平。各植被类型不同土层的土壤养分含量均随着土层的加深而减少(表2)。
从总体上看,土壤的pH值为干旱河谷地段>交错带山地森林下缘地段>交错带山地森林段;养分状况为交错带山地森林段>交错带山地森林下缘地段>干旱河谷地段。交错带山地森林段次生林的植被恢复情况最好,土壤养分含量高,其次是交错带山地森林下缘地段,干旱河谷段养分含量最低。这是因为干旱河谷受焚风气候影响,在河谷段水分条件最差,随海拔升高水分条件逐步变好[17]。在退耕还林后,随着海拔的升高,地表的植被也在逐渐地恢复,在恢复过程中林地对土壤起到的培肥作用,但这种效果并不是一直随着林分的郁闭而增加,到了山地森林段,由于青冈林为常绿阔叶林,凋落物归还少,林地中全磷、全钾以及速效钾又出现下降的趋势。处于交错带下缘的3个植被类型,土壤pH 值为灌木林>针阔混交林>白杨-旱柳混交林,土壤的养分状况为灌木林>针阔混交林>白杨-旱柳混交林。可以看出,灌木林下土壤养分含量最高,针阔混交林次之,白杨-旱柳混交人工林最低。造成差异的原因可能是由于退耕还林后虽然林木恢复较好,但林下灌丛、草本植被易受牲畜啃食、践踏,致使林内土壤板结,有机质归还量减少,而自然恢复的灌木林地,由于建群种多为沙棘(H ip pophae L.)、虎榛子(Ostryopsis david iana D ecaisne)、白刺花(Sopharo davidii(Franch.)Pavilini)等带刺植物且生长茂盛,牲畜难以进入,干扰较小,因而对土壤的改良效果较好。对人工林进行封禁管理,消除干扰因素对植被的恢复亦有重要的作用。
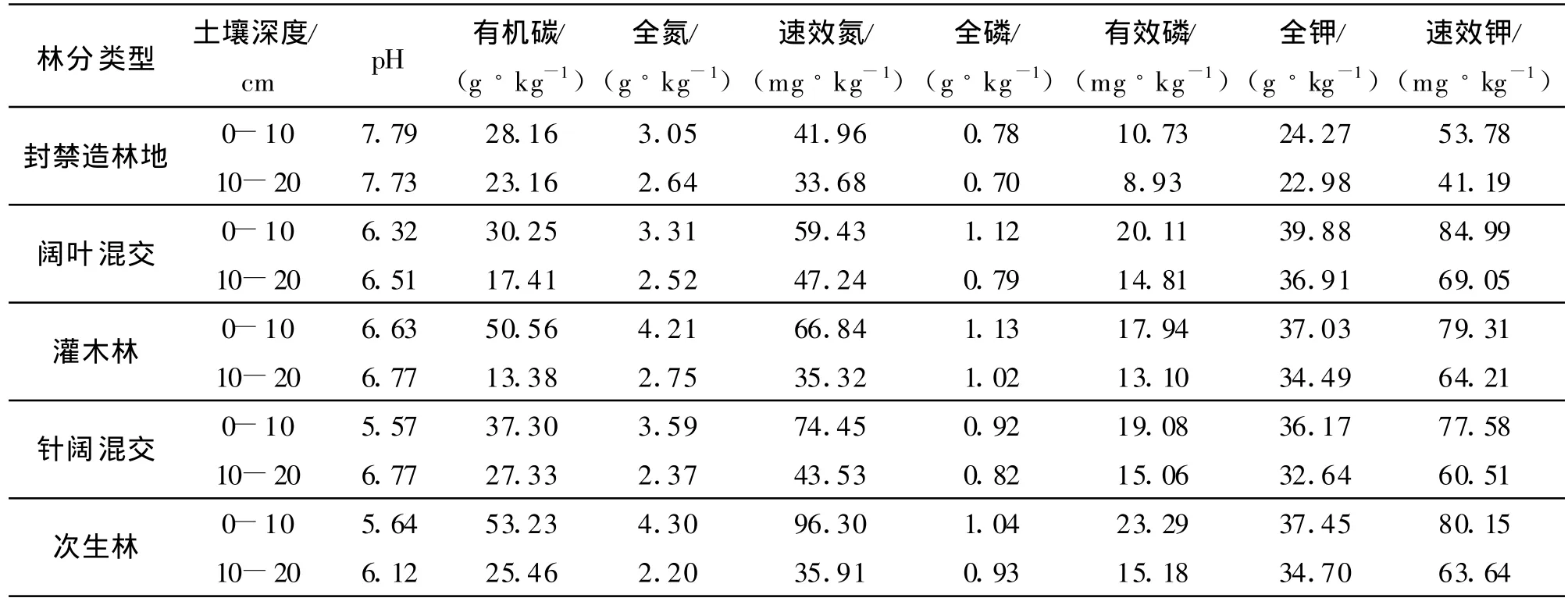
表2 不同林分类型土壤的理化性质
2.2 山地森林/干旱河谷交错带土壤微生物特征
2.2.1 土壤微生物三大类群数量分析 土壤微生物是土壤有机无机复合体的重要组成部分,其数量直接影响土壤的生物化学活性及土壤养分的组成与转化,是土壤肥力的重要指标之一[18]。各植被类型土壤的主要微生物类群的垂直分布情况如表3所示。由表3可以看出,土壤中各类微生物随着土层深度的增加而减少,究其原因是表层土壤累积有较多的有机物、养料、水分,并具有适宜的温度和氧气条件,有利于微生物的生长繁殖;并且,地表草本植物及枯枝落叶经微生物分解,随着降水向下层土壤渗透,土壤肥力由上而下降低。5种植被类型的微生物总量为灌木林>针阔混交林>次生林>白杨-旱柳混交林>封禁造林地。
从微生物的类群来看,细菌是土壤微生物总量的主要组成者,在5种植被类型中,细菌占微生物总量的91.11%~98.41%。5种植被类型的细菌与微生物总量具有相同的排列顺序,即灌木林>针阔混交林>次生林>白杨—旱柳混交林>封禁造林地。
放线菌的作用主要是分解植物和动物的某些难分解组分,形成腐殖质,把植物残体和枯落物转化为土壤有机组分。放线菌占微生物总量的1.31%~8.71%。放线菌的总量为灌木林>针阔混交林>次生林>白杨-旱柳混交林>封禁造林地。0-10 cm土层放线菌数量与放线菌总量具有相同的排列顺序;10-20 cm土层放线菌的数量为灌木林>次生林>针阔混交林>白杨-旱柳混交林>封禁造林地(表3)。
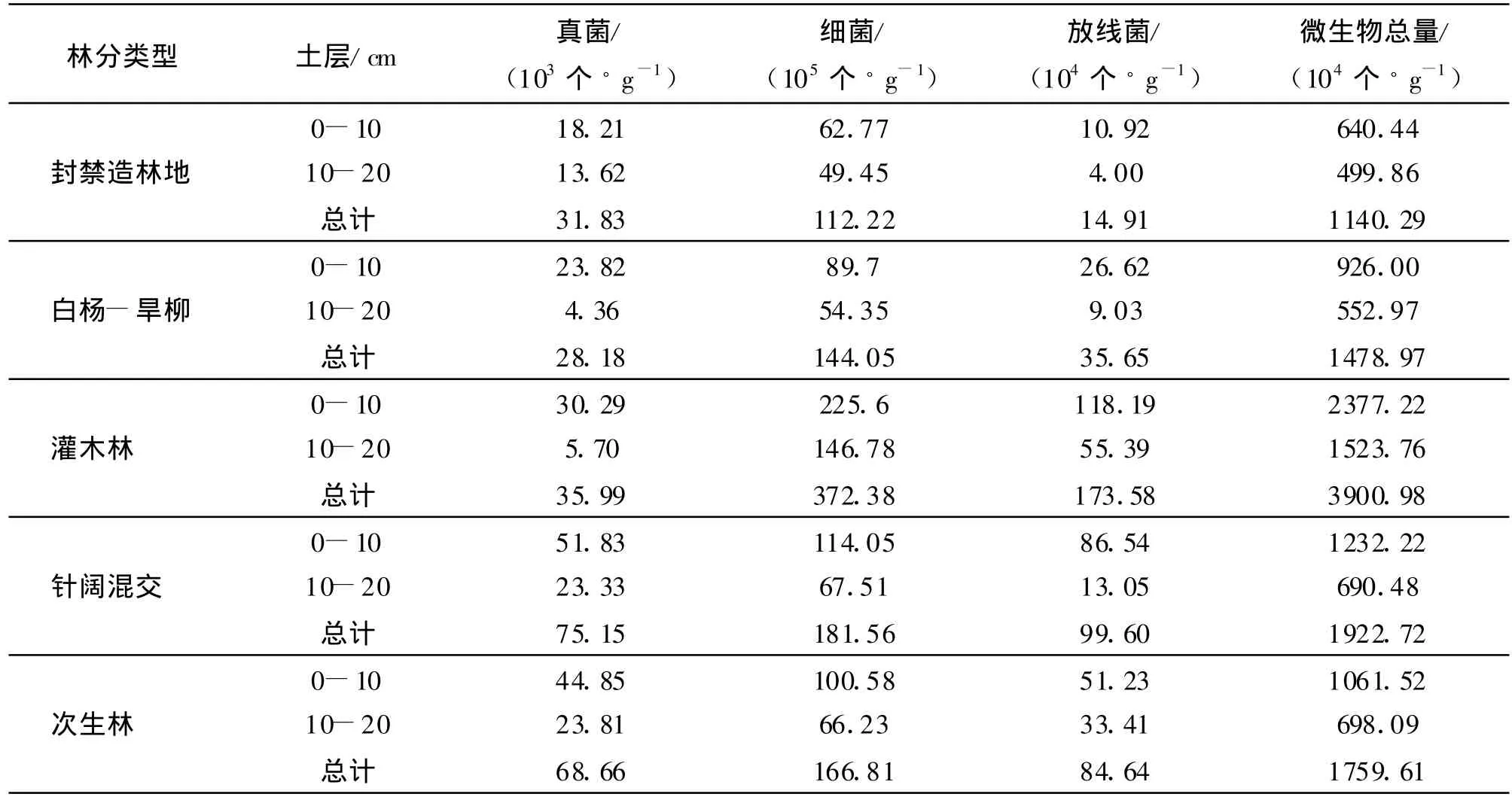
表3 3大类群土壤微生物数量
真菌积极参与有机质的分解,使凋落物中的蛋白质形成林木可直接吸收的可溶性氨基酸和铵盐等。真菌占微生物总量的0.18%~0.60%。真菌的总量为针阔混交林>次生林>灌木林>封禁造林地>白杨-旱柳混交林。0-10 cm土层真菌数量顺序为针阔混交林>次生林>灌木林>白杨-旱柳混交林>封禁造林地;10-20 cm土层真菌数量为次生林>针阔混交林>封禁造林地>灌木林>白杨-旱柳混交林。
随着海拔的升高、地表植被的恢复,不但养分得以提高,微生物总数、细菌、放线菌、真菌的数量都在交错带中达到了最高,表明随着植被的演替,枯枝落叶以及土壤理化性质的改善,微生物的生存环境也趋于稳定,使土壤更加适应了微生物的生长繁殖。随着交错带向次生林过渡,虽然土壤养分提高,但微生物的数量呈下降的趋势,是由于次生林是以常绿阔叶树青冈为主要树种,返还到地面的枯枝落叶低于落叶树种以及灌木林,符合微生物在不同林分下分布的总规律,即:灌木林>乔木林[19],阔叶纯林>针阔混交林>针叶纯林[20],落叶阔叶林﹥常绿阔叶林[6]。
2.2.2 土壤微生物生理类群数量分析 土壤微生物生理类群的垂直分布情况如表4所示。由表4可以看出,各类土壤微生物生理类群的分布与3大类群微生物呈现相似的规律。即土壤微生物主要集中在0-10 cm土层,随土层深度的增加,微生物数量减少。植被类型不同,土壤中微生物各生理类群的数量不同。山地森林/干旱河谷交错带土壤微生物各生理群数量总体表现为:氨化细菌>固氮菌>纤维素分解菌的分布特点。氨化菌、固氮菌的数量随海拔的升高先增多后减少,纤维素分解菌的数量随海拔的升高而增多。交错带山地森林下缘地段,灌木林中固氮菌、氨化菌数量最多;白杨-旱柳混交人工林中纤维素分解菌数量最多。土壤微生物各主要生理类群直接参与土壤中C、N等营养元素循环和能量流动,其数量和活性直接关系到土壤生态系统的维持和改善。固氮菌具有固定大气中氮气,增加土壤氮素的能力,对土壤的氮素补充和平衡有重大的作用;氨化细菌直接参与分解土壤中有机态N,荒地中这两类土壤微生物数量减少,降低了土壤的供N能力。荒地中纤维素分解菌数量的降低,直接影响植物残体的转化速度,使土壤中难分解的植物残体增加。由以上分析可见,荒地土壤生态系统不利于有益微生物的繁殖和活动,因而大大削弱了土壤中C,N营养元素循环速率和能量流动,这需要结合地上部分的植被修复及其它措施来达到恢复荒地土壤微生物生态系统稳定性的目的。
2.3 山地森林/干旱河谷交错带土壤微生物数量与土壤主要养分之间的关系
土壤养分含量,尤其土壤有机质是土壤微生物重要的碳源和氮源。为了探讨土壤养分与土壤微生物数量之间的关系。
从表5可以看出,山地森林/干旱河谷交错带土壤真菌数量与pH呈显著负相关关系,与全碳、速效氮呈极显著正相关关系,与全氮、有效磷显著正相关关系;细菌数量与全氮、全磷呈显著正相关关系;放线菌数量与全氮、全磷呈显著正相关关系;氨化菌数量与全碳、全磷以及全钾呈显著正相关关系;纤维素分解菌与速效氮、有效磷呈极显著正相关,与全氮呈显著正相关;固氮菌与全磷、有效钾呈极显著正相关,与全氮、有效磷、全钾呈显著正相关。本研究区域土壤微生物数量的多少均与土壤pH呈负相关关系,与土壤养分均呈正相关关系;由此可见土壤微生物数量与土壤养分含量之间存在密切的相关关系,土壤微生物数量可作为评价土壤肥力的指标。

表4 土壤微生物生理类群数量
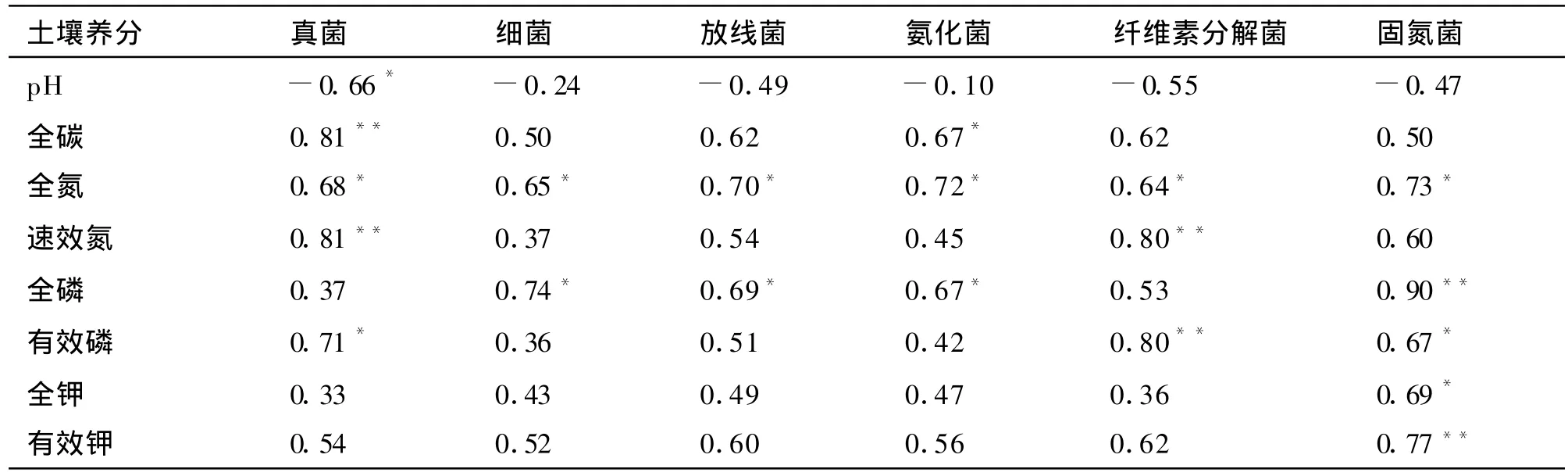
表5 土壤微生物数量与土壤主要养分的相关系数
3 结论
(1)土壤pH值为干旱河谷地段>交错带山地森林下缘地段>交错带山地森林段;养分状况为交错带山地森林段>交错带山地森林下缘地段>干旱河谷地段。随着海拔的升高,植被的恢复,土壤养分增加,pH逐渐降低。在交错带山地森林下缘地段,灌木林养分状况>针阔混交林>白杨-旱柳混交人工林。各植被类型不同土层的有机质含量、全N、全P含量随着土层的加深而减少。除荒地外,其他植被类型pH呈弱酸性并随土层的加深而增大。荒地土壤pH呈弱碱性并随土层的加深而减小。可以看出山地森林/干旱河谷交错带土壤肥力情况是高海拔优于低海拔,为干旱河谷区植被恢复从上至下逐渐推进,提高植被恢复成效提供了理论依据。
(2)土壤真菌、细菌、放线菌、氨化菌以及固氮菌的数量随海拔的升高先增多后减少,纤维素分解菌的数量随海拔的升高而增多。交错带山地森林下缘地段,灌木林中放线菌、固氮菌、氨化菌数量最多;针阔混交林中细菌、真菌数量最多;白杨-旱柳混交人工林中纤维素分解菌数量最多。各植被类型不同土层微生物的数量均随着土层的加深而减少。
(3)相关性分析表明:山地森林/干旱河谷交错带土壤微生物数量与土壤养分含量之间存在密切的相关关系,土壤微生物数量可作为评价土壤肥力的指标。
(4)次生林的各项土壤养分指标高于封禁造林地,可以看出交错带土壤条件是由海拔高处向低处逐渐变差,在干旱河谷区恢复森林植被,从上到下逐步恢复应是一个较易取得成效的途径。在交错带下缘地段,随着植被的恢复,土壤理化性状均高于封禁造林地,表明植被恢复对土壤具有明显的改良作用。至于灌木林地各项土壤养分指标高、土壤微生物数较大,则主要是由于牲畜干扰较少所致。从而也进一步表明在人为干扰较大的区域,进行封禁管理,对提高植被恢复效果具有十分重要的作用。
[1] 张荣祖.横断山区干旱河谷[M].北京:科学出版社,1992.
[2] 刘兴良,幕长龙,向成华,等.四川西部干旱河谷自然特征及植被恢复与重建途径[J].四川林业科技,2001,22(2):10-17.
[3] 王金锡.四川西部干旱河谷的生态环境与退耕还林[J].四川林业科技,2001,22(1):27-31.
[4] 关文彬,冶民生,马克明,等.岷江干旱河谷植物群落物种周转速率与环境因子的关系[J].生态学报,2004,24(11):2367-2373.
[5] 包维楷,王春明.岷江上游山地生态系统的退化机制[J].山地学报,2000,18(1):57-62.
[6] 薛立,邝立刚,陈红跃,等.不同林分土壤养分、微生物与酶活性的研究[J].土壤学报,2003,40(2):280-285.[7] 焦如珍,杨承栋,屠星南,等.杉木人工林不同发育阶段林下植被、土壤微生物、酶活性及养分的变化[J].林业科学研究,1997,10(4):373-379.
[8] 殷国兰,李梅,吴宗兴,等.岷江干旱河谷辐射松人工林土壤微生物数量的季节动态[J].四川农业大学学报,2007,25(4):410-414.
[9] 肖玲,王开运,张远彬,等.岷江冷杉根际土壤微生物对大气CO2浓度和温度升高的响应[J].应用生态学报,2006,17(5):773-777.
[10] 杨芳,王开运,杨万勤.川西亚高山不同林地土壤微生物和酶活性研究[J].内蒙古林业科技,2008,34(1):5-7.
[11] 张秀艳.川西常绿阔叶林不同恢复阶段土壤微生物区系研究[D].四川雅安:四川农业大学,2004.
[12] 杨芳.川西亚高山森林土壤微生物和酶活性分布特征研究[D].重庆北碚:西南农业大学,2004.
[13] 张秀艳,杜卫兵,张小平,等.川西植被恢复过程中的土壤微生物评价及与土壤因子的关系[J].生态环境,2007,16(5):1470-1474.
[14] 齐泽民,王开运.川西亚高山不同密度缺苞箭竹对土壤生物学特性的影响[J].水土保持学报,2007,21(4):154-158,176.
[15] 南京农业大学,中国科学院南京土壤研究所.土壤物理学[M].北京:高等教育出版社,1978.
[16] 中国科学院南京土壤研究所微生物室.土壤微生物研究法[M].北京:科学出版社,1985.
[17] 何其华,何永华,包维楷.岷江上游干旱河谷典型阳坡海拔梯度上土壤水分动态[J].应用与环境生物学报,2004,10(1):68-74.
[18] 薛立,陈红跃,邝立刚.湿地松混交林地土壤养分、微生物和酶活性的研究[J].应用生态学报,2003,14(1):157-159.
[19] 蔡艳,薛泉宏,侯琳,等.黄土高原几种乔灌木根区土壤微生物区系研究[J].陕西林业科技,2002(1):4-9.
[20] 叶镜中.森林生态学[M].哈尔滨:东北林业大学出版社,1992.
