新城疫病毒HN基因的克隆与原核表达
2010-07-04孙留霞侯喜林余丽芸
孙留霞,侯喜林,余丽芸
(黑龙江八一农垦大学,大庆 163319)
新城疫(Newcasltle disease,ND)是由新城疫病毒(Newcastle disease virus,NDV)引起的禽类一种以呼吸道、消化道黏膜出血为典型病变的急性、高度接触性传染病,被世界动物卫生组织(OIE)列为A类传染病。该病发病急、致死率高,已成为养禽业危害最大、经济损失最严重的疾病之一[1]。NDV属于副粘病毒科(Paramyxoviridae)、副粘病毒亚科(Paramyxovirinae)、禽腮腺病毒属(Avulavirus)的单股负链RNA病毒。NDV整个基因组全长约15 kb,编码6种结构蛋白:核衣壳蛋白NP、磷酸化蛋白P、RNA依赖的RNA聚合酶L、融合蛋白F、血凝素-神经氨酸酶HN、基质蛋白M[2]。HN和F为糖基化蛋白,是NDV感染细胞所必需的,为主要的免疫保护性蛋白。HN蛋白识别并吸附于细胞表面的受体,参与病毒粒子感染。F蛋白能促进病毒囊膜与宿主细胞膜融合、相邻宿主细胞之间发生融合,是构成致病力的重要成分[3]。因此F和HN蛋白的研究对防制新城疫具有重要意义。
本研究欲对NDV-Lasota株HN基因进行克隆和原核表达,通过Western blotting对表达产物的免疫原性进行研究,以期为利用HN基因研制鸡新城疫基因工程疫苗或诊断试剂提供理论依据和基础素材,并为进一步研制预防鸡新城疫基因工程疫苗奠定坚实的基础。
1 材料和方法
1.1 病毒、菌株和质粒 NDV-Lasota疫苗株购于中国兽医药品监察所;表达载体pET-30a(+)、宿主菌DH5α、BL21(DE3)由本实验室保存。
1.2 试剂 pMD18-T载体、Taq DNA聚合酶、dNTP、RNA酶抑制剂、M-MLV、限制性内切酶BamHⅠ和HindⅢ等购自Fermentas公司;Trizol Reagent购自Invitrogen公司;IPTG、胶回收和质粒提取试剂盒均为大连宝生物工程有限公产品;DAB购自AMRESCO公司;鸡源NDV标准血清购于中监所;HRP标记的羊抗鸡IgG均购自北京博奥森生物技术有限公司;其他生化试剂均为国产分析纯产品。
1.3 引物设计与合成 根据GenBank中已发表的NDV-Lasota株 HN基因全序列,利用DNAStar和Prime 5.0设计一对对引物,在上游引物中引入BamHⅠ限制性内切酶位点,在下游引物中引入HindⅢ限制性内切酶位点(酶切位点用下划线标记),预计扩增片断为1590 bp。引物由上海生物工程有限公司合成。序列如下:
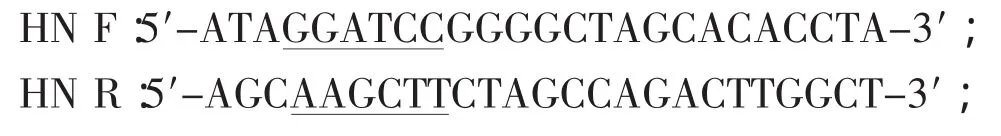
1.4 病毒的增殖 将NDV Lasota疫苗用PBS稀释后尿囊腔接种10日龄SPF鸡胚,置37℃孵育。24 h内死亡的弃掉,每天照蛋两次。无菌收集24~72 h鸡胚的尿囊液,3000 r/min离心30 min去除杂质,-20℃保存备用。
1.5 病毒RNA的提取 按照Trizol Reagent说明书提取病毒总RNA。
1.6 目的基因的扩增、克隆与测序
1.6.1 反转录合成cDNA 取5 μL提取的RNA样品,加入3 μL下游引物,3 μL无Rnase水,70℃预热 5 min,冰浴 5 min,依次加入 4 μL 5×反转录缓冲液、3 μL 10 mmol·L-1dNTPs、1 μL Rnasin(40 U/μL)、1 μL M-MLV,总体积 20 μL,42 ℃温浴 1 h。
1.6.2 PCR 扩增体系(25 μL) 10×PCR Buffer缓冲液 2.5 μL,dNTP 2 μL,上下游引物各 1 μL,Taq DNA 聚合酶 0.5 μL,模板 2 μL,加水至 25 μL。循环参数为:94 ℃ 5 min,94 ℃ 1 min,58 ℃ 1 min30 s,72℃ 1 min,共30个循环,最后72℃ 延伸10 min。
1.6.3 HN基因克隆与测序 将PCR产物纯化回收,并与pMD18-T载体连接,转化DH5α感受态细胞,用LB琼脂平板(含Amp+)筛选,提取质粒,酶切鉴定重组质粒,阳性克隆命名为pMD-HN,送上海生工测序。
1.7 重组原核表达载体的构建 用BamHⅠ和HindⅢ从pMD-HN质粒上双酶切下目的基因片断,纯化回收后与同样双酶切的pET-30a(+)载体相连,转化入BL21感受态细胞,用LB琼脂平板(含Kan+)筛选,挑取单个菌落于37℃过夜振荡培养后提取质粒,经酶切鉴定为阳性的重组质粒命名为pET-HN。
1.8 重组质粒在大肠杆菌中的表达及SDS-PAGE分析 将获得的阳性重组菌株BL21(pET-NDV/HN)按1%接种LB液体培养基(含Kan+)中,于37℃振荡培养至OD600nm为0.5~0.7时,加入IPTG至终浓度为1 mmol·L-1,诱导表达目的蛋白,并分别在 3、4 和 5 h时取1.0 ml菌液,同时收集未诱导的菌液作为阴性对照,对经转化BL21的空载体也以相同的条件诱导,作为对照。于12%的SDS-PAGE检测重组蛋白。
1.9 重组蛋白的Western blot鉴定 对诱导5 h的重组菌进行Western blot检测鉴定。蛋白样经SDSPAGE电泳后转移至硝酸纤维素膜,用含5%脱脂奶粉的PBST封闭,以鸡源NDV阳性血清为一抗,羊抗鸡IgG-HRP为二抗,DAB显色。
2 结果与分析
2.1 HN基因的扩增和克隆 RT-PCR扩增出约1590 bp左右大小的片断,与预测结果一致(图1)。回收扩增产物,克隆到pMD18-T载体中,得到重组质粒pMD-N,经BamHⅠ和HindⅢ双酶切鉴定正确(图2)。
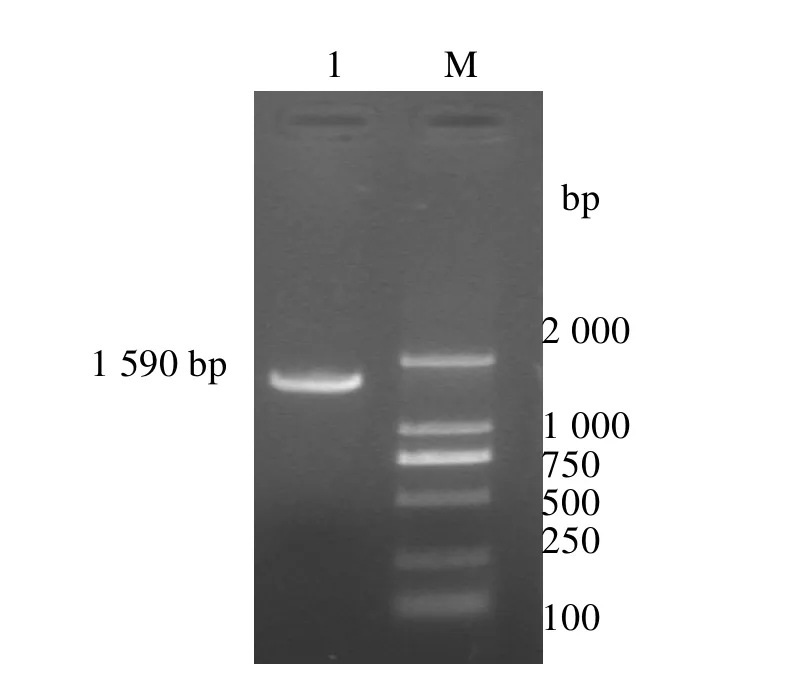
图1 HN基因RT-PCR扩增产物M.DNA标准;1.HN基因RT-PCR产物;2.阴性对照Fig.1 The amplified HN gene by RT-PCR M.DNA Marker;1.The RT-PCR product of HN;2.Negative control

图2 重组质粒pMD-HN酶切鉴定M.DNA标准;1.pMD-HN的酶切产物Fig.2 Identification of recombinant N plasmid by Restriction enzyme M.DNA Marker;1.pMD-HN digested by BamHⅠand HindⅢ
2.2 重组表达质粒的酶切鉴定 用BamHⅠ和HindⅢ双酶切鉴定重组表达质粒pET-HN,酶切结果与预期大小相符(图3)。

图3 重组表达质粒的酶切鉴定M.DNA分子质量标准;1-4.pET-HN的双酶切产物Fig.3 Identification of recombinant plasmid by restriction digestion
2.3 SDS-PAGE检测结果 IPTG诱导表达重组菌,表达产物经12%SDS-PAGE电泳、染色、脱色后,发现含重组质粒的诱导菌在64.3kDa处出现一条较粗的蛋白带,大小与融合蛋白的理论值一致,随着诱导时间的增加,其表达量随之增加,在5 h时达到最大值,而未诱导的重组质粒转化菌和空载体质粒转化菌均没有出现相同的条带(图4)。
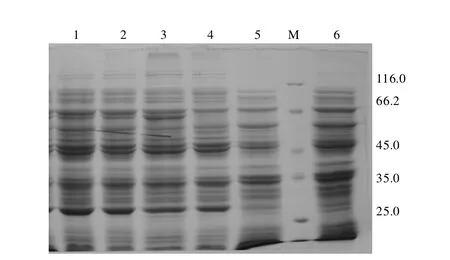
图4 IPTG诱导表达HN蛋白的SDS-PAGE分析1.诱导产物 3 h;2.诱导产物 4 h;3.诱导产物 5 h;4.重组菌诱导前;5,6.pET-30a诱导前后阴性对照;M.蛋白质分子质量标准Fig.4 SDS-PAGE of induced expression of HN protein by IPTG
2.4 重组蛋白的免疫活性鉴定 表达蛋白经PVDF膜转印后,与鸡源抗NDV阳性血清作用,再经羊抗鸡IgG/HRP二抗作用,DAB染色后,在预期位置出现明显的条带,与预计大小一致,而未诱导的重组质粒转化菌和空载体质粒转化菌均未见此特异性融合蛋白条带(如图5),说明表达的蛋白可被鸡源抗NDV阳性血清识别。

图5 重组蛋白的Western blot鉴定M:蛋白质分子质量标准;1.pET-30a;2.重组菌诱导前;3.重组HN蛋白Fig.5 Western blot analysis of the expressed product
3 讨论
HN蛋白是NDV的主要保护性抗原,在疾病感染过程中发挥重要作用,同样也是新城疫基因工程疫苗研制的首选蛋白。HN是NDV囊膜上较大纤突的糖蛋白,具有血凝素和神经氨酸酶两种活性[4]。HN借助其血凝素成分使病毒吸附到含唾液酸受体的易感细胞,这是病毒感染的第一步。而神经氨酸酶有分解病毒与细胞结合的能力,在病毒生命周期中起增加病毒粒子迁移性的作用。近年来发现,HN有促进膜融合的作用,即协助F蛋白的融合作用。它以N末端插入到病毒囊膜内,即亲水的氨基酸位于胞内,而长的疏水氨基酸跨过囊膜,形成近膜区。当NDV接触宿主细胞时,HN蛋白首先以C末端识别宿主细胞膜上的受体位点并与之结合,同时其自身构象也发生改变。当HN空间构象发生改变后,影响到相邻F蛋白的空间构象,使F蛋白N末端融合多肽释放,发挥穿膜作用,介导病毒囊膜与宿主细胞表面脂蛋白膜融合,病毒核衣壳释放到胞浆内,引起细胞病变[5,6]。
pET-30a(+)是一种十分有效的表达外源基因的原核表达载体,含T7启动子、起始密码子和转录终止信号。本试验设计引物时借助载体上的起始密码子AUG,所以pET-30-HN表达的HN蛋白分子量为64.3 kDa,而不是HN蛋白的预期大小58.3 kDa。
目前国内外学者对NDV的结构蛋白研究颇多,HN已在多种体系中表达。前人研究发现,去除HN蛋白N端的胞质区和跨膜区后的片段,这样可排除N端信号肽区和跨膜区在表达过程中的干扰,从而使去除HN蛋白N端的跨膜区和胞质区后的目的蛋白得以高水平的表达[7]。因此,本研究设计引物时,去除了N端的胞质区和跨膜区后的片段,成功表达了NDV-Lasota株的HN蛋白,为新城疫的防制和诊断方法的研究奠定了坚实的基础。
[1]殷震,刘景华.动物病毒学[M].2版.北京:科学出版社,1997.
[2]Cho S H,Kim S J,Kwon H J.et al.Genomic sequence of an antigenic variant Newcastle disease virus isolated in Korea[J].Virus Genes,2007,35(2):293-302.
[3]王忠田,王泽霖,陈博言,等.新城疫分子生物学最新研究进展[J].动物医学进展,2002,23(2):33-36.
[4]塞弗.禽病学[M].11版.北京:中国农业出版社,2005.
[5]Estevez C,King D,Seal B,et al.Evaluation of Newcastle disease virus chimeras expressing the Hemagglutinin-Neuraminidase protein of velogenic strains in the context of a mesogenic recombinant virus backbone[J].Virus Res,2007,129(2):182-190.
[6]王红宁.禽呼吸系统疾病[M].北京:中国农业出版社,2002.
[7]冉旭华,闻晓波,赵喜红,等.新城疫病毒F和HN蛋白的原核表达[J].生物技术,2008,18(3):13.
