水分胁迫下水稻幼苗多胺含量变化与抗旱性的关系
2010-06-08周小梅赵运林周朴华李小湘王淑红
周小梅,赵运林,周朴华,李小湘,王淑红
(1.湖南农业大学 生物科学技术学院,湖南 长沙 410128;2.湖南城市学院 建筑与城市规划学院,湖南 益阳413000;3.湖南省水稻研究所,湖南 长沙 410125)
多胺(polyamine,PA)广泛存在于原核生物和真核生物中,是一类低分子脂肪族含氮碱.高等植物中常见的多胺有腐胺(Put)、亚精胺(Spd)、精胺(Spm)等,它们以多聚阳离子状态存在,可与带负电荷的核酸、酶、结构蛋白以及细胞内功能基团发生作用,调节植物的生长、发育和形态建成[1-2].近年来,越来越多的的研究表明,多胺与植物对外界环境胁迫反应的关系密切.有关水分胁迫下多胺的累积与水稻抗旱性关系的研究已有报道[3-4].笔者以籼稻品种旱116(抗旱性强)、湘早籼32号(抗旱性弱)为材料,PEG6000模拟干旱对水稻幼苗进行根际胁迫处理,探讨水分胁迫下水稻幼苗多胺含量的变化与抗旱性的关系,现将结果报道如下.
1 材料与方法
1.1 材 料
旱116、湘早籼32号,在25 ℃左右的水中浸种24 h,35 ℃下催芽,选取露白一致的种子,播种于塑料杯中(每杯30粒),木村B培养液培养,于室温30 ℃自然条件下生长.
1.2 方 法
当幼苗长至二叶一心期,用含0(-0.03 MPa,处理Ⅰ)、5%(-0.10 MPa,处理Ⅱ)、10%(-0.16 MPa,处理Ⅲ)、15%(-0.33 MPa,处理Ⅳ)、20%(-0.50 MPa,处理Ⅴ)、25%(-0.77 MPa,处理Ⅵ)聚乙二醇(PEG6000)的1/2木村B培养液进行根际处理,7 d后取样测定多胺含量,各重复3次.胁迫处理后,每杯随机取幼苗20株,洗净,放入恒温干燥箱中,105 ℃杀青30 min,70 ℃烤干至恒重,分别测定幼苗地上部、根干重.
水稻幼苗多胺含量按文献[11]方法,稍加改进测定.
2 结果与分析
2.1 水分胁迫对水稻幼苗生长的影响
2.1.1 水分胁迫对水稻幼苗根系干重相对生长率的影响
水分胁迫对水稻幼苗根系生长起促进作用,随着PEG6000质量分数的增加,根系干重逐渐增加(表1),旱116根系干重的相对变化率明显大于湘早籼32号.
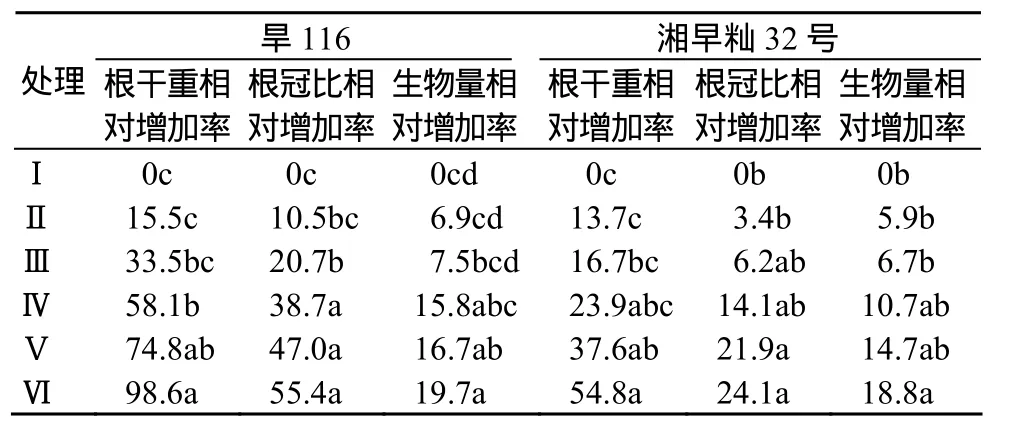
表1 水分胁迫对水稻幼苗相对生长率的影响Table 1 Effect of water stress on relative growth rate of rice seedlings%
2.1.2 水分胁迫对水稻根冠比的影响
水分胁迫促进水稻幼苗根冠比的增加,随PEG6000质量分数的增大呈上升趋势,但旱116的上升幅度明显大于抗旱性弱的湘早籼32号(表1).
2.1.3 水分胁迫对水稻总干物质积累量的影响
水分胁迫有利于水稻幼苗干物质量的积累,随着PEG6000质量分数的增加,干物质积累量呈上升趋势,旱116的干物质积累量大于湘早籼32号(表1),但增长幅度不如根系干重及根冠比明显,表明在水分胁迫下,抗旱性强的旱116的光合产物有更多的是运向根部,增强其抗旱性.
2.2 水分胁迫对水稻幼苗多胺含量的影响
2.2.1 对水稻幼苗腐胺含量的影响
从图1可看出,水分胁迫提高了水稻幼苗叶片和根系中Put含量,旱116的Put含量的上升幅度明显大于湘早籼32号,表明Put的积累与水分胁迫程度有一定相关性.湘早籼32号根系中Put含量的上升幅度明显大于叶片,而旱116在PEG6000质量分数低时叶中 Put的积累量明显大于根中,而高PEG6000质量分数时则相反(表2).
2.2.2 水分胁迫对水稻幼苗亚精胺含量的影响
从图2可看出,水分胁迫促进水稻幼苗叶片和根中Spd含量的升高,且旱116的上升幅度明显大于湘早籼32号,表明Spd含量的升高能缓解水分胁迫对水稻幼苗的伤害程度.湘早籼32号根中Spd的积累明显高于叶片,而旱116在PEG6000低质量分数时叶中增加率高于根部,而高质量分数时根中的积累量大于叶片(表2).

图1 水分胁迫对水稻幼苗叶片和根中腐胺含量的影响Fig.1 Effect of water stress on Put contents in leaves and shoots of rice seedlings
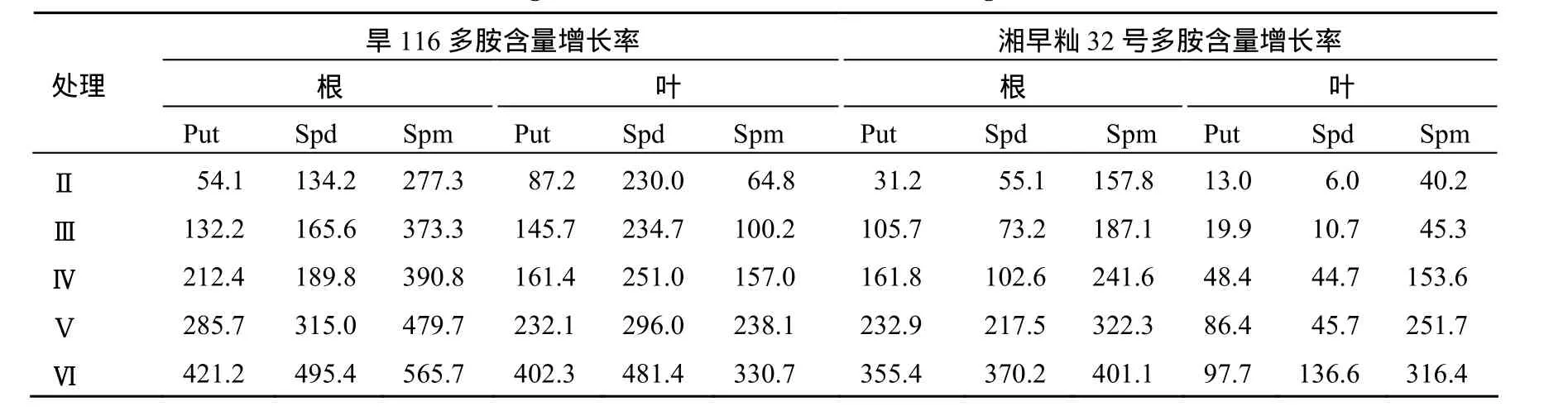
表2 水分胁迫下水稻幼苗多胺含量的相对变化率Table 2 Relative growth rate of PAs content of rice seedlings under water stress %
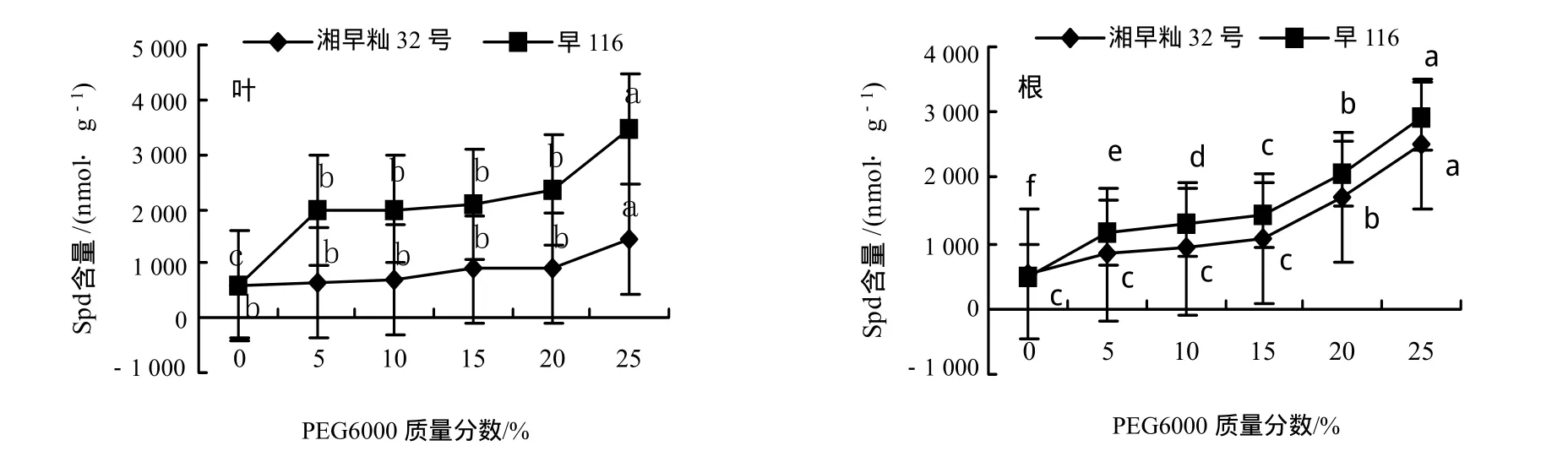
图2 水分胁迫对水稻幼苗叶片和根中亚精胺含量的影响Fig.2 Effect of water stress on Spd contents in leaves and shoots of rice seedlings
2.2.3 水分胁迫对水稻幼苗精胺含量的影响
从图3可知,水分胁迫促进水稻叶片及根中的Spm含量的上升,旱116的上升幅度明显大于湘早籼32号,表明Spm含量的上升有利于提高水稻幼苗的抗水分胁迫能力,减轻水分胁迫所带来的伤害.旱116和湘早籼32号在水分胁迫下根系中Spm的增长率明显于叶片(表2).
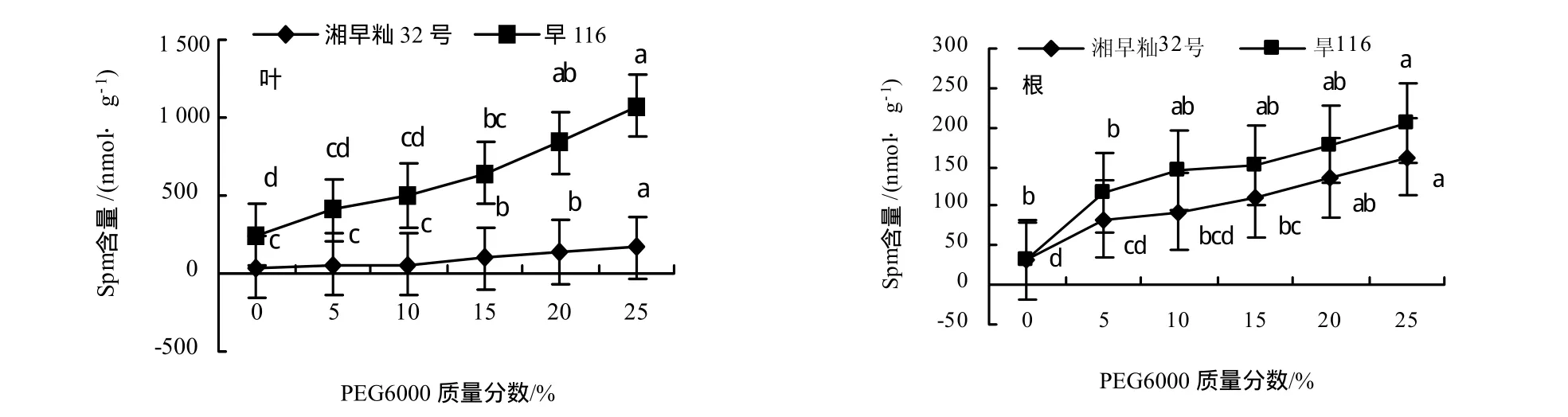
图3 水分胁迫对水稻幼苗叶片和根中精胺含量的影响Fig.3 Effect of water stress on Spm contents in leaves and shoots of rice seedlings
2.2.4 水分胁迫下水稻幼苗根部、地上部干物质的积累量与多胺的逐步回归分析
水分胁迫下,水稻幼苗根、叶中 Put、Spd和Spm的含量呈上升趋势,说明水分胁迫下Put、Spd和 Spm含量的积累与水稻幼苗抗水分胁迫能力有一定的相关性.以水分胁迫下积累的干物质量为因变量,Put、Spd和 Spm的含量为自变量进行逐步回归分析,筛选出对水分胁迫下积累的干物质量(即对抗水分胁迫能力)影响较大的多胺成分(α=0.05水平进入模型),所有方程均达到显著或极显著水平(表3).对水稻地上部而言,Put对湘早籼 32号的影响明显被引入方程,决定系数R2为0.929,达到极显著水平;对抗旱性强的旱 116,Spd的作用明显而被引入方程,决定系数R2为0.941,达到极显著水平.对根系而言,Spd和Spm对湘早籼32号影响显著被引入方程,决定系数R2为0.991,达到极显著水平,Spm的系数大于 Spd的,说明 Spm对水稻幼苗抗水分胁迫能力的影响大于 Spd;Spm对旱116根系干物质的积累影响显著被引入方程,决定系数R2为0.832,达显著水平.在4个方程当中,Spd和Spm均被2个方程引入,Put被1个方程引入,说明Put、Spd和Spm能显著提高水稻幼苗抗水分胁迫能力.

表3 水分胁迫下水稻幼苗干物质的积累与多胺成分的逐步回归分析Table 3 Regression analysis between accumulation of dry weight and PA components of rice seedlings under water stress
3 讨 论
多胺在植物的逆境胁迫中有着不同程度量的变化,各种水分胁迫都能诱导植物细胞和原生质体中Put含量的增加.但对于Put在植物抗旱中的作用的认识,仍存在分歧.番茄叶片中0.4~0.6 mol/L山梨醇水分胁迫6 h后,Put含量增加约60倍,但是Spm和Spd变化不大,而且Put的增加与ADC活性提高是平行的,但ODC活性无显著变化[5].张木清等[6]发现甘蔗叶片受到水分胁迫时多胺合成增加,特别是Put增加幅度最大,fPut/fSpd和fPut/fSpm比值均提高,抗旱性强的品种升幅明显,认为fPut含量的积累可以缓解水分胁迫对作物的伤害,提高作物的抗旱性.Gu等[7]也提出以fPut/fSpd和fPut/fSpm的比值高低作为甘蔗品种抗旱性鉴定的生理指标.Goicoechea等[8]发现,与根瘤菌共生的苜蓿因含有较高的游离态多胺,特别是 Put,故能比非共生的苜蓿较好地适应干旱胁迫.但Liu等[9]认为,水分胁迫下Put的积累不利于提高植物的抗水分胁迫能力,认为高的(fSpd + fSpm)/fPut比值有利于小麦抗水分胁迫能力的提高.Van等[10]对马铃薯的研究发现,受到干旱胁迫的马铃薯叶片内的Spd含量与马铃薯块茎的产量呈正相关,而与 Put含量无关.在本研究中,水分胁迫促进水稻幼苗叶片和根系中Put、Spd和Spm含量的上升,且抗旱性强的旱116的上升幅度大于湘早籼32号,水分胁迫下水稻幼苗根系干物质重和根冠比呈上升趋势,且旱116的根系干物质重和根冠比的上升幅度大于湘早籼32号,对根系中Put、Spd和Spm含量的变化率与根冠比的增加率进行相关性分析,相关系数R分别为:0.780、0.806和0.927,呈一定的正相关,说明不但Spd和Spm含量的上升有利于提高水稻幼苗的抗水分胁迫能力,而且Put含量的上升也能显著提高水稻幼苗的抗水分胁迫能力,此时它们可能通过稳定质膜和提高细胞内保护酶活性[12-13],以及提高 ABA含量或改变不同激素成分的含量[14-15]而增强其抗旱性.
高灿红[16]通过逐步回归分析,筛选出低温胁迫下玉米幼苗对低温冷害指标影响较大的多胺成分,根部主要与Spd有关,中胚轴主要与Put和Spd有关,胚芽鞘主要与Spd和Spm有关.在本研究中,Put和Spd分别被水稻地上部方程引用,而Spd和Spm分别被水稻根部方程引用,说明在水分胁迫下,不同多胺成分对水稻不同部位所起的作用不同.Spd和Spm对湘早籼32号根系影响显著,同时被引入方程,Spm的系数大于Spd,说明Spm对湘早籼 32号幼苗抗水分胁迫能力的影响大于Spd.Put、Spd和Spm分别被1、2和2个方程引入,进一步表明 Put、Spd和Spm是水分胁迫下的应激产物,参与了水分胁迫下的生理代谢.
根系是植株受胁迫最直接的感受器官,根系中多胺反应较叶片中更敏感[17].而在本研究中,水分胁迫时,抗旱性弱的湘早籼32号根系中Put、Spd和Spm含量上升幅度均明显大于叶片;而抗旱性强的旱116在PEG6000低质量分数时,叶片中Put、Spd含量上升幅度明显大于根系,PEG6000高质量分数时根系中 Put、Spd含量上升幅度明显大于叶片,Spm含量在整个水分胁迫过程中均是根系中的上升幅度大于叶片中,说明抗旱性有差异的水稻品种在水分胁迫时其多胺代谢存在差异,其机理尚不清楚,有待进一步探讨.
[1]Martin-Tanguy J.Metabolism and function of polyamines in plants:Recent development(new approaches)[J].Plant Growth Regul,2001,34(1):135-148.
[2]Bagni N,Torrigiani P.Polyamines:A new class of growth substances[C]//Karssen C M,Van Loon L C,Vreugdenhil D.Progress in Plant Growth Regulation.Dordrecht:Kluwer Academic Publishers,1992,264-275.
[3]杨建昌,张亚洁,张建华,等.水分胁迫下水稻剑叶中多胺含量的变化及其与抗旱性的关系[J].作物学报,2004,30(11):1069-1075.
[4]Yang J C,Zhang J H,Liu K,et al.Involvement of polyamines in the drought resistance of rice[J].Journal of Experimental Botany,2007,58(6):1545-1555.
[5]Tiburcio A F,Kaur-Sawhney R,Galston A W.Polyamine metabolism and osmotic stress[J].Plant Physiol,1981,8:375-378.
[6]张木清,陈如凯,余松烈.水分胁迫下叶片多胺代谢变化及其同抗旱性关系[J].植物生理学报,1994,22:327-332.
[7]Gu Y M G,Vigh L,Queiroz Q.Changes in polyamine and precursor content during drought induced induced adaptive morphogensis in rape[J].Francaised Actralite Botanique,1984,131:258-265.
[8]Goicoechea N,Szalai G,Antolin M C,et al.Influence of arbuscular mycorrhizae and rhizobium on free polyamines and proline levels in water-stressed alfalfa[J].Plant Physiol,1998,153(5):706-711.
[9]Liu H P,Dong B H,Zhang Y Y,et al.Relationship between osmotic stress and the levels of free,conjugated and bound polyamines in leaves of wheat seedlings[J].Plant Science,2004,166:1261-1267.
[10]Van D M A,De R J A,Van D M T,et al.Changes in free proline concentrations and polyamine levels in potato leaves during drought stress[J].South African J Sci,1998,94:347-350.
[11]Kiriakos K,Maria D,Christakis H,et al. A narrow-pore HPLC method for the identifieation and quantitation of free,eonjugated,and bound polyamines[J].Anal Bioehem,1993,214:484-489.
[12]徐仰仓,王静,刘华,等.外源精胺对小麦幼苗抗氧化酶活性的促进作用[J].植物生理学报,2001,27:349-352.
[13]周小梅.水分胁迫下水稻体内多胺代谢及其生理功能研究[D].长沙:湖南农业大学生物科学技术学院,2008.
[14]Upreti K K,Murti G S R.Effect of polyamines in the changes in endogenous hormones in pea under water stress conditions[J].Indian Journal of Plant Physiology,1999(4):1-5.
[15]聂军,郑圣先,廖育林,等.控释氮肥对水稻叶片内源激素含量及平衡的影响[J].湖南农业大学学报:自然科学版,2006,32(1):15-19.
[16]高灿红.不同耐寒型玉米种子多胺代谢的调控及对抗寒性的效应[D].杭州:浙江大学农业与生物技术学院,2006.
[17]张志新,邹志荣,张春梅,等.水分胁迫对番茄幼苗叶片和根系中多胺代谢的影响[J].西北农林科技大学学报:自然科学版,2009,37(7):97-102.
