CKMB和cTnI在缺氧诱导in vitro心肌细胞中的表达
2010-05-12甄子朋张杰
甄子朋 张杰
1临沂师范学院体育学院(山东 临沂 276005) 2四川大学华西医院卫生部移植工程与移植免疫重点实验室
CKMB和cTnI(cardiactroponin I,cTnI)均可在心肌组织中表达,二者可从功能和结构两个方面反映心脏的健康程度。CKMB是肌酸激酶(CK)的同工酶之一,剧烈活动时负责催化磷酸肌酸与ATP之间进行高能磷酸键的转换。在心肌细胞中,大多数cTnI结合于肌钙蛋白-原肌球蛋白复合物中,正常情况下,由于心肌新陈代谢的需求,约有3~8%作为代谢底物存在于胞浆内[1]。健康人血中不含或含极低量的cTnI。当心肌因各种因素发生凋亡或变性坏死时,细胞膜破损,CKMB、cTnI可弥散进入细胞间质,较早地出现在外周血中。目前cTnI已作为临床上判断心肌损伤的常规指标来使用[2,3]。
大量临床观察及研究均证实运动可以诱发心肌损伤,引起血液中CKMB及cTnI浓度升高[4]。不论临床病人还是运动员,造成心肌损伤的具体机制至今不明。Rowe[5]认为造成运动性心肌损伤的原因由两个生理性恶性循环”引起,一个是严重缺血和高儿茶酚胺,另外一个是冠状动脉痉挛和内皮损伤,它们共同作用可以引起心肌缺氧,后者可导致心肌损伤。另外,也有研究认为去甲肾上腺素可引起心肌损伤[6]。但均缺乏确切的证据。研究已证实在机体所有组织中,大部分组织氧供充足,但心肌组织即使是在正常情况下其氧含量也处于低饱和状态[7]。因此研究缺氧在心肌损伤中的地位和作用十分必要。
本研究采用RT-PCR和Western Blotting方法,观察给予单纯缺氧刺激后健康大鼠in vitro心肌细胞CKMB和cTnI表达变化,探讨表达机制,为理解缺氧诱发cTnI和CKMB表达与心脏健康之间的关系提供参考。
1 材料和方法
1.1 细胞培养
1.1.1 心肌细胞的培养
取新生Wister大鼠(四川大学华西临床医学院实验动物中心提供),在其心脏跳动时将心脏取下。小心剪取左心室组织,用冰PBS液(pH7.4)清洗干净后,将组织反复剪成0.5~1mm3的小块,加2~3滴细胞培养液,用吸管轻轻吹打后分次吸取组织块悬液,将其均匀排在75ml培养瓶(Falcon)底壁上,后加入含 150ml/L小牛血清、0.1mM 5-溴脱氧核苷 (5-Bromodeoxyuridine;Brdu)的DMEM培养液3ml,置于37℃恒温培养箱(Sanyo)中孵育30~40min后,待组织块略干能贴于瓶壁上,再缓慢、轻轻地将培养瓶翻转,使培养液浸泡组织块,继续静止培养。每天小心置于相差显微镜(Nikon)下观察,约3~4天后待细胞多数连接成片,加入0.1%胰蛋白酶(Invitrogen)1ml,放倒置显微镜(Nikon)下观察,等部分细胞呈圆形,弃胰蛋白酶液,加入含有0.1mM Brdu培养液5~10ml反复小心吹打,后1000r/min离心收集细胞,将浓度调至5×106·ml-1。
取1.5ml细胞同3ml DMEM培养液混合后,置于75ml培养瓶中继续传代培养,2~3天定量换培养液一次。供后面实验使用。
1.1.2 缺氧刺激
将细胞分成缺氧刺激组(H组)和对照组(C组)。换培养基后的第2天,H组进行如下操作:旋紧培养瓶盖,并使用无菌封口胶将瓶盖周围封闭,然后在孵箱中标准条件下缺氧孵育40分钟,后恢复正常条件继续培养。分别 于 缺 氧 刺 激 完 成 后 即 刻 、15min、30min、45min、1h、24h、48h后分别收集上清液(supernatant,SP)和细胞。将细胞用PBS稀释后,血细胞计数板计数将浓度调至5×107·ml-1,取 50μl台盼蓝染色,确定细胞的活性;其余细胞(2ml)封口胶封闭后4℃储存,备后续实验用。将上清液置于-70℃冰箱(Heraus)中备用。
1.2 生化指标的检测
1.2.1 RT-PCR二步法检测HIF-1α因子的表达
将所有收集的细胞用PBS洗一次,离心浓缩后置于DEPC处理过的EP管中,加入1ml Trizol试剂(Takara),洗头小心吹打数次后加入氯仿0.2ml(0.2ml/1mlTrizol),用力振摇 10~20s,置室温下 2~3min;8000r/min,4℃离心15min,将上层水相转移至新的1.5ml EP管中;加入0.5ml的异丙醇(0.5ml/1mlTrizol),室温沉淀 10min,8000r/min离心10min;去上清,沉淀中加入 1ml 75%乙醇溶液(DEPC水稀释),震荡混匀后,7500r/min、4℃离心5min;去上清,室温下空气挥发残存的乙醇液(约10min);加入100μl DEPC处理的水溶解RNA,吸头小心吹打溶解,后置-20℃保存;取10μl RNA做琼脂糖电泳(5%,水平电泳仪,Bio-Rad)。另取 5μl RNA稀释 20倍后在分光光度计(Bio-Rad)上测定RNA的浓度和纯度。RNA 样品的浓度 (ng/μl):A260×稀释倍数×40;RNA 样品的纯度:1.8
RT和PCR阶段反应体系的试剂配比按照试剂盒(Takara)说明书操作。内参为GAPDH,各因子引物设计采用primer5.0软件。HIF-1α引物序列为F:5’-TCAAAGTCGGACAGCCTCA-3’;R:5’-CCCTGCAGTAGGTTTCTCT-3’;GAPDH 引物序列为 F:5’-GAAGGTGAAGGTCGGAGTC-3’;R:5’-GAAGATGGTGATGGGATTTC-3’。反应条件为:94℃变性 30s→53℃褪火 30s→72℃延伸 4min。上述循环次数:30次。琼脂糖(2.0%)电泳鉴定表达情况。
1.2.2 细胞总蛋白的提取
将收获的细胞以0.1M pH7.4的冷PBS洗涤3次,加入600μl RIPA裂解液,同时按照100∶1加入鸡尾酒蛋白酶抑制剂(Calbiochem)和磷酸酶抑制剂(Invitrogen),4℃振荡 30min后超声 10s,以 13000r/min离心 30min后,用BCA蛋白检测试剂盒(Pierce)测定提取蛋白浓度,存于-70℃冰箱(Heraus)中备用。
1.2.3 Western Blotting检测CKMB和cTnI的表达
配制分离胶和积层胶(15%)。加等量加样buffer,后将样品100℃中煮沸5min。上样进行SDS-PAGE电泳(垂直电泳仪,Bio-Rad)。开始电压为80V,当样品跑出浓缩胶,进入分离胶后,将电压调至120V。等电泳完成,将蛋白转膜至PVDF膜(Millipore)上。后用5%脱脂奶粉于室温封闭2h。后将一抗(羊抗,Pierce)用5%脱脂奶粉稀释,把含有蛋白的PVDF膜置于其中,4℃震荡孵育过夜。第2天,用 TBST(10mmol/L的 Tris碱、150mmol/L氯化钠、0.1%Tween 20、pH 8.0)室温摇洗 3×10~15 分钟。后用HRP标记的二抗(Pierce)继续孵育 2h(室温),TBST摇洗。后加Super Signal West Dura Extended Duration Substrate检测免疫反应性。最后在暗室内进行自显影(按照试剂盒说明)。使用扫描仪(联想,LS5260U)扫描胶片、得图,后使用Image Pro-Plus(3.0版本)进行IOD分析。
1.3 统计学分析
2 结果
2.1 HIF-1α表达变化
缺氧刺激后即刻,胞内即有HIF-1α表达,在30min达峰值后逐渐下降,48h后消失。

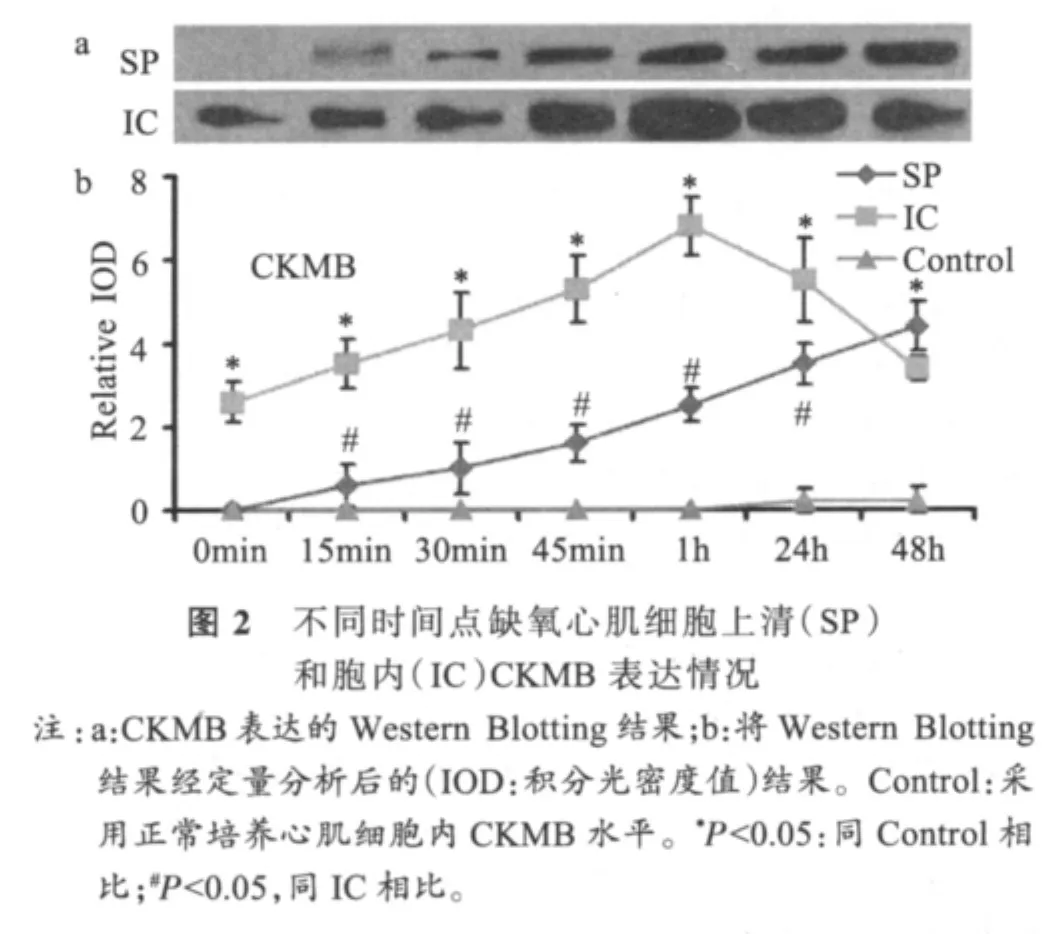
2.2 CKMB的时相性变化
图2显示,缺氧刺激后,上清中和胞内(intracellular,IC)均有CKMB表达。上清中CKMB升高趋势较平缓,一直持续到实验结束。缺氧后0min胞内CKMB水平显著升高(P<0.05),1h达峰值,后明显下降。缺氧刺激后,胞内与上清中CKMB浓度的显著性差异从15min起一直持续到24h(P<0.05)。缺氧心肌细胞内CKMB浓度一直显著高于对照细胞内浓度(P<0.05)。
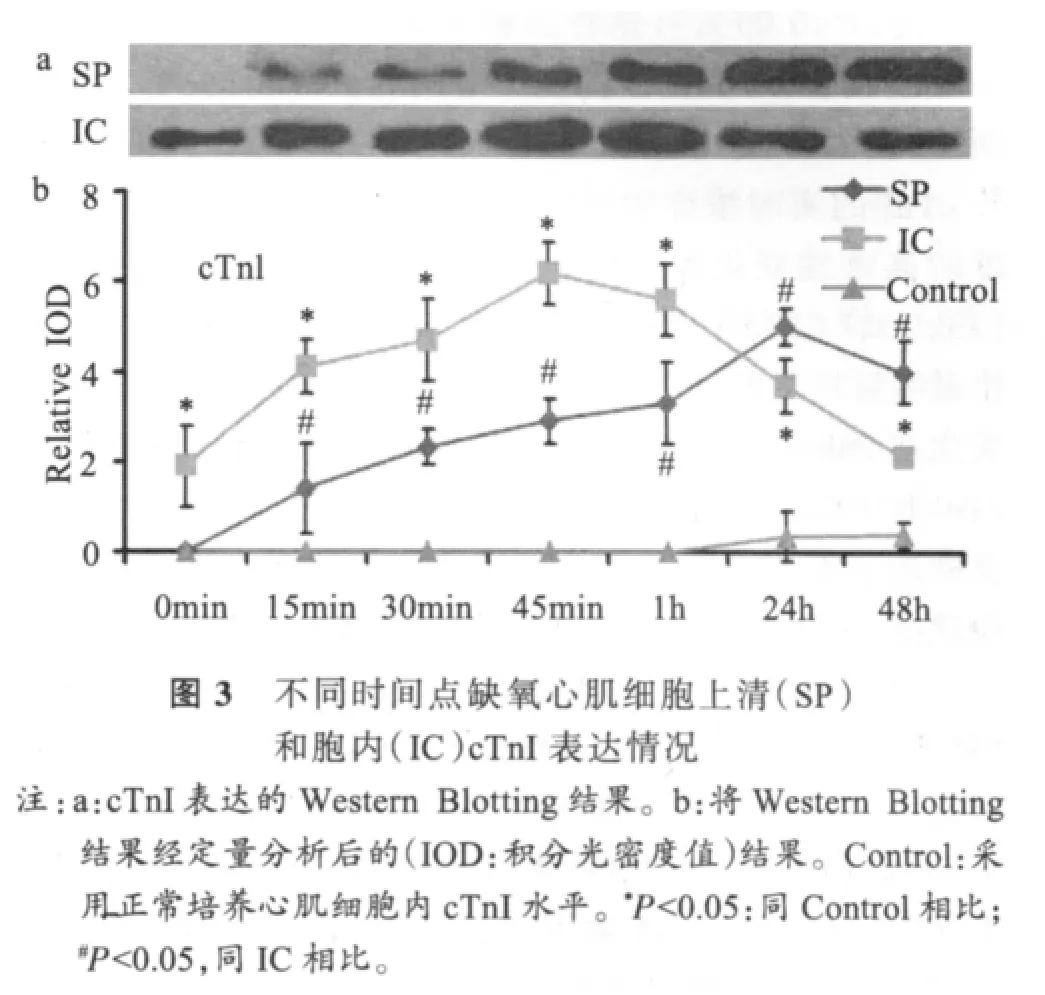
2.3 cTnI的时相性变化
图3显示,缺氧刺激后15min,上清中cTnI浓度急剧升高,且随时间延续其升高趋势亦延续,24h后达峰值,48h后仍然高表达。胞内则持续表达cTnI,随时间延续逐渐升高,45min后达到峰值,后明显下降。在实验的48h内,SP和IC呈近似同步性表达。缺氧心肌细胞内与上清中cTnI浓度除0min时间点外,其余时间点均存在显著性差异(P<0.05)。缺氧心肌胞内cTnI浓度在所有检测时间点均显著高于对照心肌细胞内浓度(P<0.05)。
3 讨论
本实验中,缺氧诱导培养的in vitro心肌细胞胞内CKMB表达量显著升高(P<0.05),这些增加的CKMB可能主要是在缺氧刺激之下细胞内有氧能量代谢受阻,而无氧供能系统代偿性增强的结果。本研究发现,in vitro心肌细胞在40min缺氧刺激后,胞内和培养上清中CKMB和cTnI均呈升高趋势。胞内CKMB峰值出现在1h,而上清峰值在实验结束时仍未检测到,推测40min缺氧刺激引起心肌细胞内能量链的连续性失调,使心肌收缩所需ATP短缺,激活了CK,代偿心肌能量供应。胞内这种生理功能性反应一直持续到1h,后减轻,48h后已低于即刻时间点。24h后上清CKMB浓度显著低于胞内(P<0.05),而48h后上清浓度高于胞内。这种关系变化意味着1h后胞内反应以功能性恢复过程占主导,而胞外CKMB浓度变化的反应要延迟。推测这可能与CKMB分子量较大(85kD),溢出细胞的速度受限有关。
心肌受缺氧刺激可以通过不同信号传导途径引起细胞的复杂反应。一方面缺氧使线粒体内ATP酶内的电子传递链阻滞,大量电子漏出,不仅ATP产生受阻,而且产生大量自由基[8],后者可氧化胞膜的脂质双分子层,破坏胞膜完整性。另一方面可以启动细胞的保护过程,促进细胞因子表达。血管内皮细胞生长因子(VEGF)表达增加,促进血管生成(angiogenesis),同时提高血管通透性(permeability)。由于心肌内氧含量始终处于低氧状态[7],因此VEGF表达升高的生理效应可以提高心肌组织的供氧效率,保护心肌。另外,缺氧可通过mTor途径激活HIF-1基因,HIF-1表达一方面表明胞内处于缺氧状态,同时也强化了细胞的生存保护性生理反应。缺氧也可通过AMPK途径使蛋白质和糖原的合成增加,胞内CKMB升高可能也是心肌细胞对于缺氧刺激的生理反应的结果之一[9]。故推测本研究中上清CKMB就是胞膜被自由基破坏后,从胞浆内渗出的结果。
本研究证实,缺氧40min后胞内cTnI表达量显著高于上清(P<0.05),且这种趋势从缺氧后即刻一直持续到1h后。而在1h到48h的三个时间点上,二者关系发生了倒转,在24h和48h,胞内cTnI浓度均低于上清。细胞内、外cTnI的实时浓度的时间差比CKMB明显,且内、外浓度的连续性变化呈近似同步性。推测这与cTnI分子量23kD)较CKMB小有关。就二者而言,胞内对缺氧刺激发生最强反应的时间点在45min~1h,而上清中cTnI的峰值发生在24h后,CKMB则不明显。针对此结果,似乎上清中TnI比CKMB更能实时反映胞内情况。但考虑到CKMB反映的心脏的功能状态,而cTnI仅是体现心脏结构的健康状况,因此在实际应用中,建议二者兼顾,更为全面。
正常心肌细胞内“游离”cTnI是细肌丝新陈代谢的储备库”[1]。心肌是高活性组织,新陈代谢较旺盛,因此对胞浆内cTnI浓度的变化敏感。推测在施加40min缺氧后,心肌细胞胞膜完整性被自由基破坏,胞内“储备库”中部分“游离”的cTnI溢出胞膜,使细肌丝新陈代谢平衡被破坏。为维持细肌丝结构完整性,新陈代谢需求压力、HIF-1α参与的mTor胞内信号途径及缺氧刺激介导的p38MAPK信号传导途径的合力作用,激活了胞内cTnI相关基因的表达,促使胞内 cTnI表达量升高[10,11]。cTnI基因应激性表达启动,胞内45min后达到峰值,而上清中的峰值出现在24h后。两个峰值出现的时间差意味着大量的cTnI驻留在胞内,供心肌细胞在缺氧应激时的生理功能所需,可能包括心肌收缩成分的更新、修复。这可能也是包括心肌细胞在内所有结构应激时代偿性反应的一部分。结合cTnI在胞内的存在形式和百分比这些事实,推测本研究检测到的上清cTnI可能是以胞浆中“游离”形式的cTnI为主,但不能确定是否含有结构性cTnI成分。
本实验结果尚难对单纯40min缺氧刺激对于in itro心肌细胞细肌丝的完整性是否会造成损伤得出明确结论。由本研究推测,马拉松及铁人三项等运动员赛后CKMB和cTnI一过性升高的部分原因是由于心肌细胞膜被自由基破坏并从心肌组织渗出。24~48h后,氧供有效恢复,自由基被清除,胞膜完整性得以修复,这些指标即恢复正常。而对于心梗病人,其原因要复杂得多。其缺氧部位心肌组织长时间缺氧,推测其胞膜也不能得到及时的修复,cTnI漏出也不能及时终止,因此即使是新生的cTnI也会漏出,最终其收缩功能得不到有效的恢复。从这个意义上讲,无论临床病人还是长时间大强度运动项目的运动员心脏,最有效的保护措施之一就是在运动中和运动后尽可能保持、恢复心脏的氧供。
对于从事长时间大强度运动项目的运动员心脏健康的检测和评价,目前尚无统一标准,而借鉴临床的近似标准的弊端也显而易见。如何确定CKMB和cTnI的变化同运动员心脏健康程度的关系仍然是疑问。本研究仅就单纯缺氧刺激对in vitro心肌细胞结构和功能的影响进行了研究,但临床及运动相关的cTnI和CKMB升高是复杂反应的结果,能对心肌的结构和功能造成重要影响的不仅仅有缺氧,年龄、血脂、生化因子的变化、能源物质供应等因素均应考虑在内[4,12],我们将在后续的 in vivo研究中就相关问题进行研究、探讨。
4 小结
单纯缺氧刺激可导致in vitro心肌细胞释放CKMB和cTnI。推测是在缺氧刺激后,生存应激反应启动,胞内功能和结构相关基因被激活,表达量升高的结果;由于缺氧诱导心肌细胞膜的完整性被破坏或者是通透性增大等原因,使二者从胞内漏出。但上述推论尚需研究予以证实。
[1]Korff S,Katus HA,Giannitsis E.Differential diagnosis of elevated troponins.Heart,2006,92(7):987-993.
[2]Antman E,Bassand JP,Klein W,et al.Myocardial infarction redefined--a consensus document of the Joint European Society of Cardiology/American College of Cardiology committee for the redefinition of myocardial infarction.J Am Coll Cardiol,2000,36(3):959-969.
[3]Ilva T,Eriksson S,Lund J,et al.Improved early risk stratification and diagnosis of myocardial infarction,using a novel troponin I assay concept.Eur J Clin Invest,2005,35(2):112-116.
[4]Shave R,George K,Gaze D.The influence of exercise upon cardiac biomarkers:a practical guide for clinicians and scientists.Curr Med Chem,2007,14(13):1427-1436.
[5]Rowe WJ.Extraordinary unremitting endurance exercise and permanent injury to normal heart.Lancet,1992,340(9):712-714.
[6]Kondo T,Ogawa Y,Sugiyama S,et al.Mechanism of isoproterenolinduced myocardial damage.Cardiovasc Res,1987,21(4):248-254.
[7]Henquell L,Odoroff CL,Honig CR.Coronary intercapillary distance during growth:relation to PtO2and aerobic capacity.Am J Physiol,1976,231(6):1852-1859.
[8]张勇,文立,聂金雷.运动性疲劳的线粒体分子机理研究,线粒体质子跨膜势能与运动性内源自由基生成的关系.中国运动医学杂志,2000,19(4):346-348.
[9]Brancaccio P,Maffulli N,Limongelli FM.Creatine kinase mon-itoring in sport medicine.Br Med Bull,2007,81-82(1):209-230.
[10]Yang Q,Guan KL.Expanding mTOR signaling.Cell Res,2007,17(8):666-681.
[11]Roux PP,Blenis J.ERK and p38 MAPK-activated protein kinases:a family of protein kinases with diverse biological functions.Microbiol Mol Biol Rev,2004,68(2):320-344.
[12]Febbraio MA,Steensberg A,Keller C,et al.Glucose ingestion attenuates interleukin-6 release from contracting skeletal muscle in humans.J Physiol,2003,549(2):607-612.
