OPN和BMP15基因与大长杂交一代母猪产仔数的关系
2010-05-09孙敬礼李大全刘贤侠
孙敬礼,武 磊,黄 涛,李大全,刘贤侠
(1.石河子大学动物科技学院,新疆 石河子 832003;2.新疆克拉玛依瑞恒畜牧开发有限责任公司,新疆 克拉玛依 834000)
母猪的产仔数是一个重要的经济性状,产仔数的高低直接影响到猪场的经济效益。因此,有效地提高猪的产仔数,特别是提高猪的产活仔数,一直是科学研究和养猪生产的主要任务之一。近年来,随着分子生物学技术和现代繁育理论的不断发展,从基因水平对猪高繁殖力机制进行探讨已成为一研究主题。骨桥蛋白(osteopontin,OPN)是一种含精氨酸-甘氨酸-天冬氨酸的分泌型糖基化磷蛋白,广泛存在于乳汁、子宫、胎盘等组织和体液中,具有多种生物学活性。鉴于其在动物繁殖中的重要功能。,近年来,它成为在猪繁殖性状的重要候选基因之一[1]。骨形成蛋白 15(bone morphogenetic protein-15,BMP15)是由卵母细胞分泌的一种生长因子,对早期卵泡的生长、分化和排卵起重要作用;许多研究表明BMP15可能具有促进卵泡发育,防止黄体早熟的生物学功能[2]。目前,许多学者都将其作为控制产仔数的候选基因加以研究。本文主要分析大长杂交一代母猪OPN和BMP15不同基因型以及合并基因型对经产母猪的产仔数差异,为利用DNA标记辅助选择方法提高母猪综合繁殖性能提供理论依据。
1 材料和方法
1.1 试验动物
产仔2胎以上且记录完备的大长杂交一代母猪样本119头,采自瑞恒公司猪场。前腔静脉采血,EDTA抗凝。
1.2 DNA提取
采用苯酚-氯仿-异戊醇抽提方法提取DNA,置于-20 ℃冰箱中保存备用。
1.3 引物设计
本研究所用引物分别参照Knoll等[3]、候振平等[4]所发表的引物序列由上海生工生物技术公司合成引物。OPN-F:5 -TCACCGATTTCCCCACCGACAC-3 ;OPN-R :5 -TGGCTGCGGGTTTCCACACTG-3 。BMP15-F :5 -GATGCGTCTGCCACCTACAT -3 ;BMP15-R :5 - CCGGAACTCAAGAATCTCAC -3 。

1.4 PCR扩增
OPN、FSHβ和ESR基因采用相同的反应体系:10×Buffer 2.5,μL,dNTP2μL,上、 下 游 引 物 各0.5 μL(10μmol/L),50 ng/μL 模 板 DNA 1μL,Taq 酶 1U,Mgcl2 2 μL,加双蒸水至 25 μL。
OPN基因的PCR扩增反应条件:94 ℃变性4 min,35 次循环(94 ℃,40 s;60 ℃,1 min;72 ℃,1.1 min),72 ℃延伸 10 min。
BMP15基因的PCR扩增反应条件:94℃变性4 min, 35 次 循 环(94 ℃,30 s;58 ℃,30 s;72 ℃,30s),72 ℃延伸 10 min。
1.5 多态性检测
OPN基因由片段插入造成,琼脂糖电泳检测PCR产物即可见多态性,PCR产物经1.5%的琼脂糖凝胶电泳检测,在凝胶上显示为 3种带型 :1295 bp、1295/990 bp和 990 bp, 分 别 为 AA、AB和 BB型。BMP15基 因采 用 PCR-RFLP 进 行 分 析 :8.5μL PCR 产 物 加 入0.5 μL Spe Ⅰ及 1.5 μL 10×Buffer,37℃水浴 4 h,酶切产物用2%的琼脂糖电泳检测分型:显示1条带(503 bp)的为 AA 型,显示 2 条带(277 bp、226 bp)的为 BB 基因型,显示 3 条带(503 bp、277 bp、226 bp)为 AB 基因型。
1.6 数据处理
整理基因型数据,计算所测猪群体OPN和BMP15基因不同基因型的基因型频率、基因频率,用SPSS13.0分析产仔数在各基因型间的差异显著性,数据用平均数±标准差表示。
2 结果与分析
2.1 基因型频率和基因频率
根据OPN、BMP15基因型检出个数,计算基因型频率和基因频率,见表1。OPN基因AA型频率最大,BB型频率最小,AB在两者之间,且A等位基因占优势,分别为0.673和0.327;BMP15基因BB型较少,AA型最多,A基因频率为0.613,为优势等位基因。
2.2 OPN和BMP15基因与产仔数关系
根据大长杂交一代猪群中OPN基因和BMP15基因各基因型及其合并基因型检出个数和对应的经产产仔数资料,计算OPN基因和BMP15基因与产仔数关系。
从表2可以看出,BB型产仔数比AA型高1.717头,产活仔数比AA型高1.223头;不同基因型间产仔数表现出BB>AB>AA的趋势。AA型与BB型对猪的产仔数差异极显著(P<0.01),但对猪的产活仔数差异显著(0.01<P<0.05)。
由表3可以看出,AA和AB型个体对猪的产仔数和产活仔数均高于BB型个体,AA型与AB型、BB型对猪的产仔数和产活仔数差异显著(0.01<P<0.05),AB型与BB型差异不显著(P>0.05)。
从表4可以看出,BMP15和OPN合并基因型中BBBB合并基因型对猪的产仔数、产活仔数最高,分别为12.750和11.500;BBAA合并基因型产仔数、产活仔数最低,产仔数只和BBAA、ABAA合并基因型差异不显著(P>0.05),和其他合并基因型差异极显著(P<0.01)。
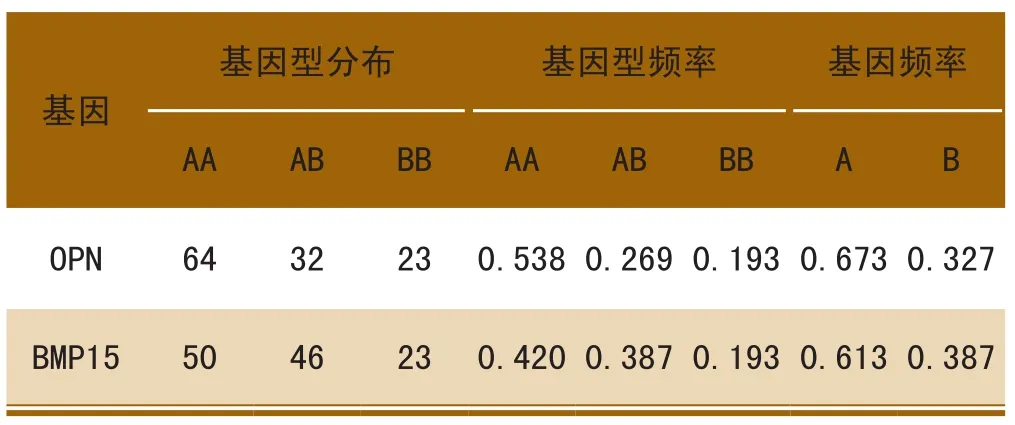
表1 OPN和BMP15基因的基因型分布和频率
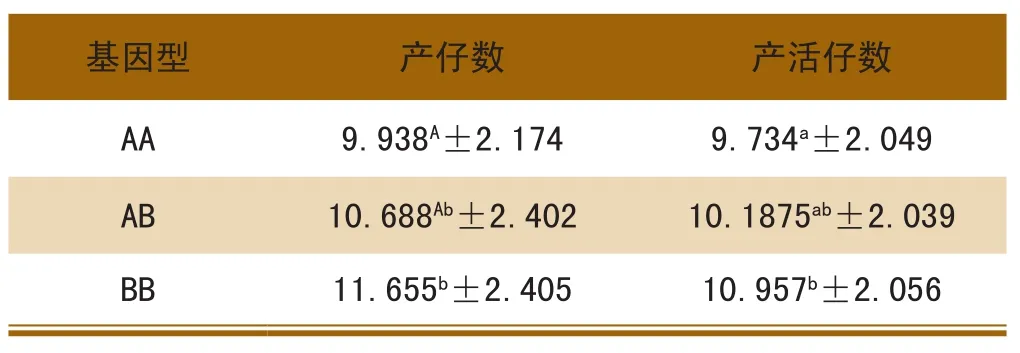
表2 OPN基因与产仔数的关系

表3 BMP15基因与产仔数的关系
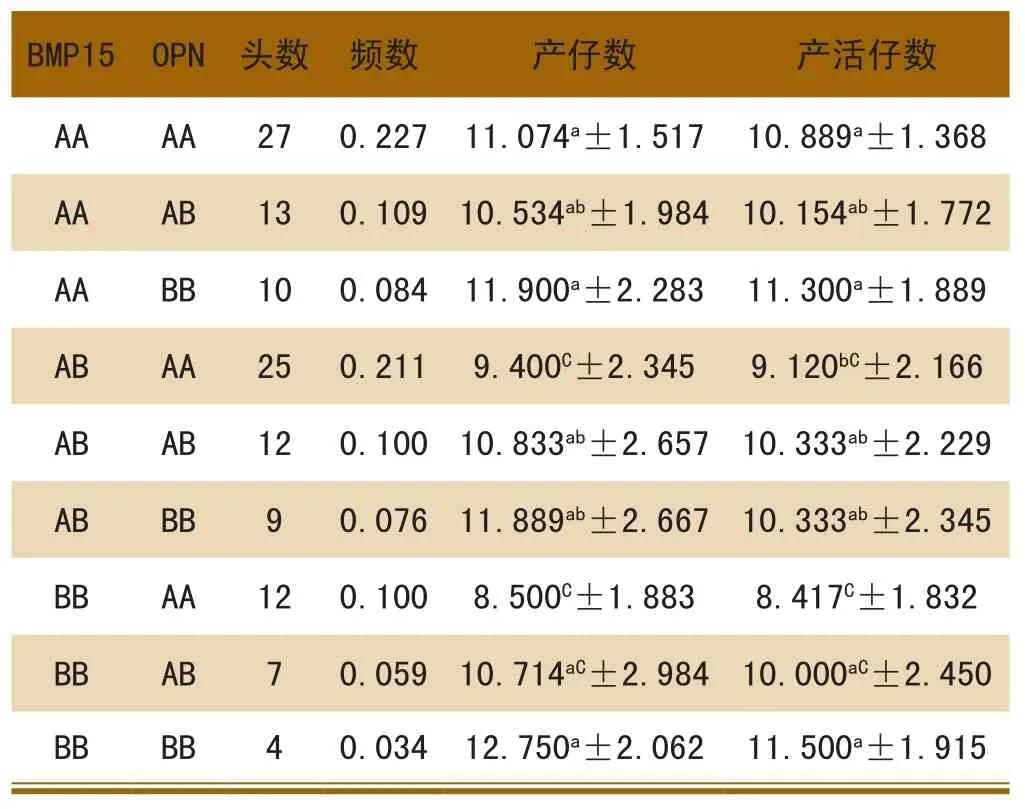
表4 BMP15和OPN合并基因型与产仔数的关系
3 讨论
OPN基因的多态性非常丰富,有研究表明,骨调素基因启动子区存在9个(TG)n双核苷酸的交替重复序列,是骨调素基因转录速率的主要区域[5]。Liaw[6]在约克夏、杜洛克、盘克夏和蓝瑞斯母猪群体中共发现27种基因型;Southwood发现了13个等位基因。此外,在OPN基因的内含子上也存在多态,Knoll[3]研究发现猪OPN基因的第6内含子上有305 bp的缺失,并发现长白猪、皮特兰猪上A等位基因占优势(0.57和0.82),而大白猪和杜洛克猪的B等位基因占优势(0.74和0.84)。而本实验结果表明A和B两个等位基因在大长杂交一代母猪群体中均有分布,且A等位基因占优势;就基因型分布而言,AA基因型频率最大,BB基因型频率最小,AB在两者之间;从表2可以看出,BB和AB型个体对猪的产仔数高于AA型个体。这与罗仍卓么等[5]的研究指出的在长白和大白猪种中,OPN优良基因型为BB型是一致的。
关于BMP15基因位点对繁殖性能的影响已有相关报道,许多研究表明在绵羊中BMP15基因突变杂合体能引起排卵数的增加。储明星等[7]研究表明BMP15FecXG突变杂合基因型小尾寒羊平均产羔数比野生纯合基因型多0.62只或0.55只。侯振平等[4]研究发现,AA型母猪群体中,二花脸、梅山猪的产仔数和产活仔数极显著高于BB型母猪群体;AB型梅山猪群,头胎产仔数和产活仔数极显著高于AA型和BB型母猪群体。本研究中,BMP15基因的3种基因型个体都检测到了。另外,本研究表明,大长杂交一代猪的不同基因型母猪的总产仔数和产活仔数间差异显著,从表3可以看出,AA产活仔数比BB高1.335头;不同基因型间产仔数表现出AA>AB>BB的趋势。这与侯振平等[4]和吴井生等[6]的研究指出的在梅山和大白猪种中,AA型母猪的产仔数和产活仔数显著高于BB型母猪群体是一致的。
从表4可以看出,BMP15和OPN合并基因型中AABB和BBBB合并基因型产仔数、产活仔数均高于其它合并基因型,BBBB合并基因型最高,理论上应是AABB合并基因型的最高,在本次研究中却是BBBB合并基因型最高,这可能因为BBBB合并基因型的样本量太少有关,头数只有4个,其与产仔数的关系需进一步加大样本量。BBAA合并基因型产仔数、产活仔数最低;这与理论上是一致的。因此,对于OPN基因而言,B等位基因均可视为增加产仔数的优势基因。这和单独应用OPN基因型的分析结果[8]一致。对于BMP15基因而言,A等位基因可视为增加产仔数的优势基因。
从本次检测结果来看,被检测的大长杂交一代猪BMP15基因的有利基因A频率较高,为0.613;但是OPN基因的有利基因B频率较低,仅为0.327;在随后的育种生产实践中,我们将根据基因检测结果,结合产仔记录和生产性能,选择OPN为BB基因型和BMP15为AA基因型的母猪和后备猪,改善生产母猪繁殖力的遗传基础,加快繁殖力性状的遗传进展,增加母猪的窝产仔数,以提高生产水平和经济效益。

[1] Yin Li, He Z Q and Zhong G.Osteopontin, it′ s receptor and the reproduction (J). Progressin Obstetrics and Gynecology, 2005,14(3):239 ~ 240
[2] 吴井生,朱孟玲.骨形成蛋白15基因BcuI多态性对猪产仔性能的影响[J].畜牧与兽医,2009,41(3):19.
[3] Knoll A, Stratil A, Cepica S and Dvorak J. Lengthpolymorphism in an intron of the porcine osteopontin (SPP1)gene is caused by the presence or absence of a SINE (PRE-1)element (J). Animal Genetics, 1999, 30:466
[4] 候振平,印玉龙,等.猪骨形成蛋白15基因编码区序列的克隆及测序研究[J].广西农业科学,2006,25(2):91.
[5] 罗仍卓么,王立贤,孙世铎.猪OPN基因与繁殖性状的关联分析[J].农业生物科技学报,2008,16(3):412-416.
[6] Liaw R B, Chang H L, Lai Y Y, Liu C T and Wu M C. Influence of sows' osteopontin alleles to their litter performance (J).Journal Chinese Society of Animal Science, 2000, 29(Suppl.): 131
[7] 储明星,桑林华,王金玉,等.小尾寒羊高繁殖力候选基因BMP15和GDF9的研究[J].遗传学报,2005,32(1):38-45.
[8] 孟庆利,刘铁铮.猪骨调素基因多态性对产仔性状的影响[J].家畜生态学报,2005,26(3):13-16.
