黄芩素对人胰腺癌PANC-1细胞增殖和运动能力的影响
2010-04-06任学群李宜雄
任学群,李宜雄
(1.河南大学淮河医院 普通外科,河南 开封 475000;2.中南大学湘雅医院 普通外科,湖南 长沙 410008)
黄芩素对人胰腺癌PANC-1细胞增殖和运动能力的影响
任学群1,李宜雄2*
(1.河南大学淮河医院 普通外科,河南 开封 475000;2.中南大学湘雅医院 普通外科,湖南 长沙 410008)
目的:探讨黄芩素对人胰腺癌PANC-1细胞增殖、运动能力的影响及其机制。方法:体外培养人胰腺癌PANC-1细胞,运用MT T法、划痕愈合实验(Wound healing assay)、扫描电镜和 Transwell实验等方法,观察黄芩素对人胰腺癌PANC-1细胞增殖、伪足形成、侵袭和迁移能力的影响,并运用Western印迹分析、RT-PCR检测黄芩素干预后肿瘤细胞Ezrin蛋白、磷酸化Ezrin蛋白及Ezrin mRNA表达的变化。结果:黄芩素呈时间和剂量依赖性抑制PANC-1细胞增殖,使肿瘤细胞伪足形成显著减少,细胞的运动侵袭能力下降。Western印迹分析和 RTPCR结果显示,黄芩素明显下调Ezrin-mRNA和Ezrin蛋白表达及其磷酸化活化。结论:黄芩素抑制肿瘤细胞恶性增殖及运动侵袭能力,可能与其抑制肿瘤细胞Ezrin表达及活化有关。
Ezrin;胰腺癌;黄芩素
近年来,广泛存在于自然界植物中的黄酮类化合物因低毒且具有多种生物活性而逐渐引起学者们的关注,尤其是其大多数具有较强的抗肿瘤活性,正日益成为抗肿瘤药物研究的热点。黄芩素是黄芩干燥根中提取的黄酮类化合物。研究表明:黄芩素是选择性12-脂氧酶(12-lipoxygenase,12-LOX)抑制剂,具有诱导肿瘤细胞周期停滞、促进肿瘤细胞凋亡、抑制肿瘤新生血管生成、增强肿瘤细胞对化疗药物的敏感性等作用,具有广谱的抗肿瘤生物活性。此外,黄芩素还具有抗肿瘤侵袭转移的作用,但机制不明。
作者前期的研究显示,Ezrin在胰腺癌细胞恶性增殖和运动转移过程中具有重要作用,且下调其表达可显著抑制胰腺癌细胞的恶性增殖及侵袭迁移能力。本研究中,作者以体外培养的人胰腺癌PANC-1细胞株为研究对象,观察黄芩素对肿瘤细胞增殖、运动侵袭能力及Ezrin表达或活性的影响,探讨黄芩素抗肿瘤侵袭和转移的可能作用机制。
1 材料和方法
1.1 细胞培养
人胰腺导管癌PANC-1细胞株购自中国科学院上海细胞生物研究所,用含体积分数为10%胎牛血清、10 mmol/L 的HEPES、10万 U/L 的青霉素、100mg/L的链霉素和4 μmol/L谷氨酰胺的DEME培养基置于37℃、体积分数为5%CO2、饱和湿度的二氧化碳培养箱中培养。
1.2 四甲基偶氮唑蓝(MTT)法测细胞增殖
用无水乙醇将黄芩素(购自南京替斯艾么中药研究所,HPLC≥98%)稀释成浓度梯度为:1.0、2.0、4.0、8.0、16.0、32.0 μmol/L,共 6 个梯度。取对数生长期的细胞常规消化制成单细胞悬液,并调整细胞密度为1×104/mL,接种于 96 孔板中,每孔培养液 200μL。培养24 h待细胞贴壁后,加入上述各浓度的黄芩素(总体积200 μ L/孔),乙醇对照组加等体积无水乙醇,空白组加等体积培养液。继续培养 24 h后,开始MTT反应,570nm波长下检测每孔吸光值(OD)。连续测72 h,每个时间点每组设6个复孔,按下列公式计算细胞生长抑制率:细胞生长抑制率(IR)=(1-实验组平均OD值/对照组平均OD值)×100%。并以抑制率为纵轴,分别以浓度对数、不同浓度梯度和时间为横轴,绘制直线回归曲线、48 h浓度依赖曲线和时间-浓度量效曲线,用直线回归法推算黄芩素作用的50%抑制浓度(IC50),为下面的实验提供依据。
1.3 Western印迹分析
加小剂量黄芩素(5、10μmol/L)培养 48h 后 ,提取细胞总蛋白,经聚丙烯酰胺凝胶电泳后转移至PVDF膜上,用含50 g/L的脱脂奶粉的封闭液封闭1h,转入4℃封闭过夜。加入1:1000稀释的鼠抗人Ezrin单克隆抗体、鼠抗人磷酸化Ezrin单克隆抗体(美国Upstate公司)或鼠抗人-actin单克隆抗体(美国Abcam公司),4℃孵育过夜;洗一抗,加1:3000稀释的辣根过氧化物酶标记的IgG(美国Sant-Cruz公司),室温孵育2 h,洗二抗,ECL显影,观察,摄片。应用Quantity one软件进行定量分析。
1.4 细胞总RNA提取及RT-PCR
对数生长期细胞用无血清培养基培养12 h,分别加入5.0、10.0 μmol/L黄芩素继续培养48 h后,收集细胞,Trizol(美国 Invitrogen公司)法提取细胞总RNA,逆转录出cDNA后进行PCR反应,琼脂糖凝胶电泳,凝胶成像系统拍照并进行电泳条带分析。各步骤按说明书进行。引物设计和合成委托上海吉凯公司完成,引物终浓度为2 μmol/L。引物序列:Ezrin上游:5′-CTCATCCAGGACATCACCCA-3′,下游 :5′-TCACTCCAAGGAAAGCCAAT-3′,扩增片段长度为 450 bp;内参 β-actin 上游 :5′-GACAGGATGCAGAAGGAGAT-3′,下游 :5′-TGTGTGGACTTGGGAGAGGACT-3′,扩增片段长度为550 bp。PCR扩增条件:94℃预变性10min,94℃变性45 s,52℃退火30 s,72℃延伸30 s,35个循环;72℃延伸10min。Ezrin mRNA的相对表达量以Ezrin RT-PCR产物电泳条带的光密度值与β-actin的光密度值的比值表示。
1.5 流式细胞仪(FCM)检测细胞周期和凋亡
细胞用分别含 5.0、10.0 μmol/L黄芩素培养基培养48 h,收集各组细胞1×106个,体积分数为70%冰乙醇固定,碘化丙锭(PI)染色,流式细胞仪检测细胞周期和细胞凋亡率。
1.6 划痕愈合实验(Wound healing assay)
细胞按2×105/孔接种于6孔板,加入含DMSO的无血清DMEM培养3 h,使细胞呈现出单层贴壁生长状态。用200 μ L的tip头给6孔板中的细胞划痕,然后改用含2 g/L G418和体积分数为10%FBS且分别含5.0 μmol/L 、10.0 μmol/L 黄芩素的DMEM,在体积分数为5%CO2、37℃培养箱中继续培养48 h。分别于划痕后0、24 h、48 h给细胞拍照,观察划痕愈合能力,每组细胞实验3个样本,重复3次。
1.7 Transwell实验检测细胞的运动侵袭能力
采用孔径为8 μ m的24孔Borden小室(美国BD公司),加1:30稀释的Matrigel胶(美国BD公司)100 μ L制作人工基底膜。用含体积分数为10%FBS且分别含 5.0 μmol/L 、10.0 μmol/L 黄芩素的 DMEM培养基重悬细胞,调整细胞数,上室中加入2×105个细胞,总体积200 μ L,下室中加入600 μ L 含体积分数为10%FBS的DMEM培养基。37℃、体积分数为5%CO2培养箱中孵育40 h后结晶紫染色,200倍光镜下随机选择5个视野计数穿膜细胞数,取其平均值作为穿过人工基底膜的细胞数量。
1.8 扫描电子显微镜观察细胞伪足
用含体积分数为10%FBS且分别含5.0 μmol/L、10.0 μmol/L黄芩素的DMEM 培养基培养48 h后,收集细胞铺片,体积分数为 2.5%戊二醛固定。按照扫描电镜标本处理程序进行标本制备,在扫描电镜(HITACHI R-250)下观察细胞伪足形态和数量。在2000倍视野下随机选取5个细胞作伪足计数,取其平均值作为该细胞的伪足数,并进行统计学分析。
1.9 统计学处理
应用SPSS 10.0统计软件进行统计学分析。所有计量资料采用均数±标准差()表示,重复测量的计量资料采用重复测量的方差分析进行检验,其他资料用单因素方差分析或t检验,所有检验均为双侧检验,并以α=0.05作为检验水准。
2 结果
2.1 黄芩素对PANC-1细胞增殖的影响
黄芩素对PANC-1细胞增殖具有明显的抑制作用,同一时相各剂量组间、同一剂量组不同时相间比较差异均有显著性(P<0.01),呈明显的剂量和时间依赖性,见表1、图1。
表1 黄芩素对PANC-1细胞增殖的影响(%,,n=6)

表1 黄芩素对PANC-1细胞增殖的影响(%,,n=6)
黄芩素/μmol˚ L-1抑制率/%24h 48h 72h 000011.51±0.62 2.33±0.91 8.17±1.8525.33±1.35 10.23±2.44 24.67±4.4348.82±1.31 13.82±4.74 31.43±3.79821.64±1.83 43.52±6.81 76.13±1.8016 37.99±1.99 53.63±5.34 85.54±7.0632 51.35±4.38 82.63±7.79 91.88±8.43
选择48 h时,以细胞增殖抑制率为纵坐标,以黄芩素浓度的对数为横坐标,应用Excel 2300做出半对数曲线,建立的回归线性方程为:y=32.704x+10.567,r为0.9003(P=0.000),建立的回归方程有意义。并根据此方程计算出48 h时黄芩素对PANC-1细胞增殖的IC 50是16.0694 μmol/L,见图2,3。

图1 黄芩素对PANC-1细胞时间-剂量增殖抑制曲线(B1~ B6 分别为黄芩素 1、2、4、8、16、32μmol/L)
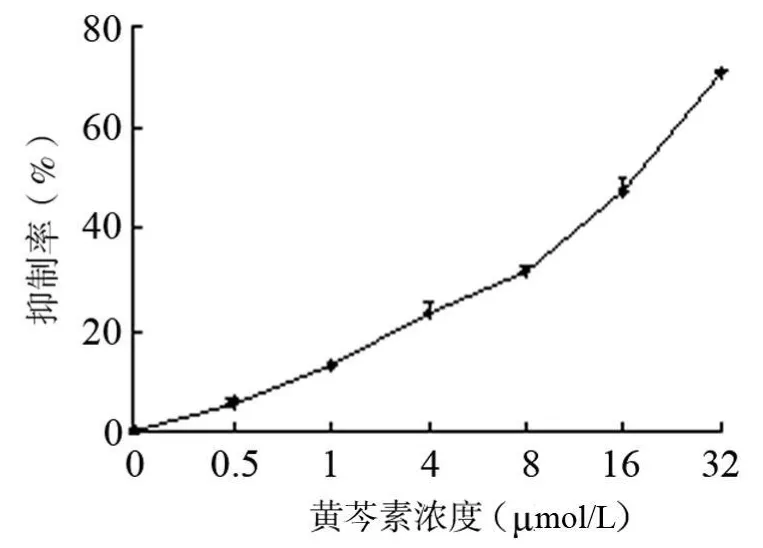
图2 黄芩素48h对PANC-1细胞剂量增殖抑制曲线

图3 48h黄芩素对PANC-1细胞增殖抑制作用的半对数曲线
2.2 Western blot检测黄芩素对 PANC-1细胞Ezrin和磷酸化Ezrin蛋白表达的影响
根据MTT比色法检测结果而求出的48h时黄芩素对PANC-1细胞增殖的半数抑制浓度(IC50),我们用小剂量黄芩素(5、10 μmol/L)处理48h后 ,分别提取细胞总蛋白用于Western blot检测。结果显示:5μmol/L黄芩素培养 48h后,Ezrin蛋白表达降低,但与对照组比较,差异无显著性(P>0.05),而磷酸化Ezrin蛋白表达明显降低,与对照组比较,差异有显著性(t=3.430,P=0.006);10 μmol/L黄芩素干预后,Ezrin和磷酸化Ezrin蛋白表达的抑制作用明显增强,与对照组比较,差异均有显著性(t=2.735,P=0.021;t=3.997,P=0.003),见图4、表2。

图4 Western blot检测黄芩素对PANC-1细胞Ezrin和磷酸化Ezrin蛋白表达的影响
表2 黄芩素处理后Ezrin和磷酸化Ezrin蛋白相对含量比较(n=6)

表2 黄芩素处理后Ezrin和磷酸化Ezrin蛋白相对含量比较(n=6)
与对照组比较*P<0.01;※P<0.05。
组别 Ezrin Phos-Ezrin黄芩素处理组/μmol˚ L-1 5.0 0.75±0.180.45±0.10※10.0 0.55±0.19*0.35±0.10*对照组 0.88±0.150.67±0.16
2.3 RT-PCR检测黄芩素对 PANC-1细胞 Ezrin mRNA表达的影响
5 μmol/L黄芩素培养48 h后,Ezrin mRNA表达即有抑制作用,但与对照组比较,差异无显著性(t=2.169,P=0.055),而 10 μmol/L 黄 芩素对PANC-1细胞Ezrin mRNA表达的抑制作用明显增强,与对照组比较,差异有统计学意义(t=5.423,P=0.000),见图 5 、表 3。
2.4 黄芩素对PANC-1细胞爬行能力的影响
划痕愈合实验检测PANC-1细胞爬行能力(闭合宽度/原宽度,%),结果显示:24 h后,对照组细胞形态较正常,呈长梭形及多角形,透亮,划痕区域可见相当数量的类梭形细胞生长,少数细胞呈圆型,不透亮,划痕基本上达到愈合;黄芩素5 μmol/L处理组细胞生长缓慢,呈短梭形,划痕区域有细胞爬行,残留有划痕存在,与对照组比较,差异有统计学意义(t=8.951,P=0.000);黄芩素10 μmol/L处理组细胞部分死亡,残存细胞呈短梭形,透亮,划痕区域只有几乎无细胞爬行,与对照组及 5 μmol/L处理组比较,差异有统计学意义(t=34.294,29.791;P=0.000,0.000),见表4。

图5 RT-PCR检测Ezrin mRNA在PANC-1细胞中的表达(PCR 产物大小:Ezrin450 bp、β-actin 550 bp)
表3 黄芩素处理后 Ezrin mRNA相对含量比较(,n=6)

表3 黄芩素处理后 Ezrin mRNA相对含量比较(,n=6)
*与对照组比较 P<0.01。
组别 Ezrin mRNA黄芩素处理组/μmol˚L-1 5.0 0.50±0.0910.0 0.30±0.09*对照组 0.63±0.12
表4 黄芩素对 PANC-1细胞爬行能力的影响(%,,n=8)

表4 黄芩素对 PANC-1细胞爬行能力的影响(%,,n=8)
*与对照组比较 P<0.01,△与黄芩素 5μmol处理组比较 P<0.01。
组别 划痕时间/h 024黄芩素处理组/μmol˚L-1 5.0 0.0±0.0 64.5±4.5﹡10.0 0.0±0.0 10.5±2.5﹡△对照组 0.0±0.0 88.0±10.3
2.5 Transwell实验观察黄芩素对PANC-1细胞运动侵袭能力的影响
黄芩素 10 μmol/L处理组 PANC-1细胞的迁移/侵袭能力均明显下降,穿膜细胞数明显减少,与对照组及5 μmol/L处理组比较,差异均有统计学意义(t=4.163/7.620,4.223/4.745;P=0.003/0.000,0.003/0.001),见表 5。
2.6 扫描电子显微镜下观察黄芩素对PANC-1细胞伪足的形态和数量的影响
在2000倍视野下可见对照组细胞形态饱满,有较多的片状伪足和大量的线状伪足,伪足比较伸展,而黄芩素10 μmol/L处理组细胞体积缩小,伪足变短且数量减少,与对照组及5 μmol/L处理组比较差异均有统计学意义(t=8.374,5.578;P=0.000,0.001),见表6。
表5 黄芩素对PANC-1细胞迁移/侵袭能力的影响(平均细胞数/视野,n=5)

表5 黄芩素对PANC-1细胞迁移/侵袭能力的影响(平均细胞数/视野,n=5)
*与对照组及黄芩素5μmol处理组比较 P<0.01。
组别 穿膜细胞数无Matrige 包被Matrigel黄芩素处理组/μmol˚ L-1 5.0 192.6±36.8 158.0±34.910.0 103.6±29.4* 70.0±22.4*对照组 232.0±62.4 181.8±24.0
表6 黄芩素对PANC-1细胞伪足形成的影响(n=5)

表6 黄芩素对PANC-1细胞伪足形成的影响(n=5)
*与对照组及黄芩素5μmol/L处理组比较 P<0.01。
组别 细胞伪足数黄芩素处理组/μmol˚ L-1 5.0 15.2±2.410.0 8.6±1.1*#对照组 18.0±2.2
3 讨论
筛选有效的抗肿瘤侵袭转移药物,对提高胰腺癌治疗效果、改善其恶劣预后,无疑具有重要的理论意义和临床应用价值。本研究结果显示,黄芩素具有较强的抗肿瘤细胞恶性增殖作用,且呈明显的时间和剂量依赖性。有研究[1]显示,黄芩素还具有逆转肿瘤细胞的多药耐药作用,对化疗药物具有协同增效效应。而绝大多数胰腺癌细胞对目前临床应用的化疗药物耐药,提示黄芩素对抗胰腺癌治疗具有潜在应用价值。
目前的研究[1-9]认为,黄芩素抗肿瘤作用机制主要集中于以下几个方面:①影响花生四烯酸系统代谢途径。花生四烯酸是多种生物活性物质的前体,虽然在生理状态下水平很低,但当细胞膜受到各种刺激(如生长因子、化学因子等)或磷脂酶功能异常时,细胞膜磷脂在磷脂酶A2和磷脂酶C的作用下分解释放出游离的花生四烯酸。花生四烯酸至少可经过三条代谢途径,形成具有生物活性的二十碳衍生物,即细胞色素P-450途径、环氧合酶途径和脂氧酶途径。脂氧酶家族成员大致分为四大类,即5-LOXs、血小板型12-LOXs、12/15-LOXs和表皮型 LOXs,其中,血小板型12-LOX将花生四烯酸转化为12-羟廿碳四烯酸(12-hydroxyeicosatetraenoicacid,12-HETE)。研究表明12-HETE是一种重要的信号分子,可通过 NF-κ B、ERK1/2、PKC 、PI3 k 和 SrcK 等多条信号传导通路,参与对肿瘤细胞增殖、运动、侵袭、血管生成和凋亡等的调节。大量研究证实黄芩素是选择性12-LOX抑制剂,可通过抑制12-LOX表达和活性,抑制12-HETE的生成,从而发挥其抗肿瘤活性。②影响细胞周期多种调控因子的表达或活性,诱导肿瘤细胞增殖周期停滞。③影响凋亡相关基因的表达或活性、激活线粒体死亡通路等,诱导肿瘤细胞凋亡。④抗肿瘤新生血管生成。黄芩素不仅可显著抑制血管内皮细胞增殖、迁移、芽突形成及形成血管样结构,而且,还可抑制VEGF的表达,抑制肿瘤血管生成。此外,黄芩素还具有抗肿瘤侵袭转移的作用。体外研究[5-9]显示,用黄芩素处理的肿瘤细胞,丝状突起明显减少,细胞密度降低,黏附性、运动和迁移能力显著降低。动物试验[8-9]也显示,原位注射种植用黄芩素处理的人高转移性肿瘤细胞,可显著减少肿瘤肺转移。本研究结果亦显示,黄芩素可显著减少肿瘤细胞伪足,降低肿瘤细胞的运动侵袭能力。
在前面的研究中,我们发现膜-细胞骨架连接蛋白Ezrin在胰腺癌细胞的侵袭转移过程中具有重要作用,且下调其表达可显著抑制胰腺癌细胞的恶性增殖及侵袭迁移能力。本研究结果显示:以低于IC50剂量的小剂量黄芩素即可下调Ezrin mRNA和Ezrin蛋白表达,且随剂量增加而抑制作用显著增强。提示下调Ezrin蛋白表达可能是黄芩素抑制肿瘤侵袭转移的可能机制之一。
研究[10-11]表明,通常情况下,Ezrin C端与N端能够发生分子内的相互作用,形成头尾相连的折叠状态,或与ERM家族其他成员结合,从而以失活状态存在于胞浆中,而Ezrin蛋白的磷酸化激活是Ezrin发挥其对细胞骨架动态循环调控功能的必需条件。本研究结果还显示:相比较而言,黄芩素对Ezrin蛋白的磷酸化抑制作用更强,提示黄芩素还可通过抑制Ezrin蛋白的磷酸化而发挥抗肿瘤细胞侵袭和转移作用。
本研究结果表明,黄芩素不仅具有较强的抑制胰腺癌细胞恶性增殖作用,而且还可抑制胰腺癌细胞膜-细胞骨架连接蛋白Ezrin的表达及其活化,可能具有抗胰腺癌侵袭转移的潜在应用价值。而且,作为一种传统中药,黄芩素还具有毒、副作用小的特点,价格低廉,药源充足,很有开发前景。
[1]徐珊,罗莉,朱利群,等.4′-甲醚-黄芩素对绒癌耐药细胞株多药耐药性的逆转作用研究[J].生物化学与生物物理进展,2006,33(11):1061~1073.
[2]Chen FL,Wang XZ,Li JY,et al.12-lipoxygenase induces apoptosis of human gastric cancer AGS cells via the ERK1/2 signal pathway[J].Dig Dis Sci,2008,53(1):181-187.
[3]Chao JI,Su WC,Liu HF.Baicalein induces cancer cell death and proliferation retardation by the inhibition of CDC2 kinase and survivin associated with opposite role of p38 mitogen-activated protein kinase and AKT[J].Mol Cancer Ther,2007,6(11):3039-3048.
[4]Lee HZ,Leung HW,Lai M Y,et al.Baicalein induced cell cycle arrest and apoptosis in human lung squamous carcinoma CH27 cells[J].Anticancer Res,2005,25(2A):959-964.
[5]Günther S,Ruhe C,Derikitomg,et al.Polyphenols prevent cell shedding from mouse mammary cancer spheroids and inhibit cancer cell invasion in confrontation cultures derived from embryonic stem cells[J].Cancer Lett,2007,250(1):25-35.
[6]Ding XZ,Kuszynski CA,El-Metwally TH,et al.Lipoxygenase inhibition induced apoptosis,morphological changes,and carbonic anhydrase expression in human pancreatic cancer cells[J].Biochem Biophys Res Commun,1999,266(2):392-399.
[7]Nie D,Nemeth J,Qiao Y,et al.Increased metastatic potential in human prostate carcinoma cells by overexpression of arachidonate 12-lipoxygenase[J].Clin Exp Metastasis,2003,20(7):657-663.
[8]Timá r J,Rásó E,Döme B,et al.Expression,subcellular localization and putative function of platelet-type 12-lipoxygenase in human prostate cancer cell lines of different metastatic potential[J].Int J Cancer,2000,87(1):37-43.
[9]Pidgeon GP,Tang K,Cai YL,et al.Overexpression of platelet-type 12-lipoxygenase promotes tumor cell survival by enhancing αvβ3and αvβ5integrin expression[J].Cancer Res,2003,63(14):4258-4267.
[10]Bretscher A,Edwards K,Fehon RG.ERM proteins and merlin:integrators at the cell cortex[J].Nat Revmol Cell Biol,2002,3(8):586-599.
[11]Fievet BT,Gautreau A,Roy C,et al.Phosphoinositide binding and phosphorylation act sequentially in the activation mechanism of Ezrin[J].J Cell Biol,2004,164(5):653-659.
[责任编辑 姬 荷]
Effect and mechanism of baicalein on human pancreatic cancer PANC-1 cell proliferation and metastasis
REN Xue-qun1,LI Yi-xiong2*(1.Department of General Surgery,Huaihe Hospital,Henan University,Kaifeng,Henan 475000,China;2.Department of General Surgery,Xiangya Hospital,Central South University,Changsha,Hunan 410008,China)
Objective:To investigate the effect and its probable mechanism of baicalein on human pancreatic cancer PANC-1 cell proliferation and metastasis.Methods:Human pancreatic cancer PANC-1 cells were cultured in vitro.Cell proliferation was detected by MTT method,cell pseudopodia was observed by scanning electron microscopy,the migration and invasion ability of cells were detected by wound healing assay and trans well assay,expressions of Ezrin and phosphate-Ezrin protein were detected by western blotting,and Ezrin mRNA was detected by reverse transcriptase-polymerase chain reaction(RTPCR)in PANC-1 cells treated by baicalein.Results:Baicalein suppressed PANC-1 cell proliferation in a time and dose dependent manner.Treated with baicalein,the number of pseudopodia per cells,the migration andinvasion ability of PANC-1 cells decreased significantly.Reverse transcriptase-polymerase chain reaction(RT-PCR)and Western blotting results revealed that baicalein notably regulated down Ezrin expression at both mRNA and protein levels,and inhabited its phosphorylation as well.Conclusion:Baicalein can inhibit human pancreatic cancer PANC-1 cell proliferation,invasion and metastasis,which may be the results of down-regulation the expression level of Ezrin mRNA and Ezrin protein as well as its phosphorylation.
Ezrin;Pancreatic Adenocarcinoma;Baicalein
R735.9
A
1672-7606(2010)03-0180-06
2010-03-10
任学群(1964-),男,河南正阳人,博士,教授,主任医师,从事肝胆胰外科方面的研究工作。
*通讯作者:李宜雄(1959-),男,湖南嘉禾人,教授,主任医师,博士生导师,从事胰肝外科的临床工作及相关基础研究。
