卫星搭载对白三叶光合特性的影响
2010-03-31王健刘荣堂张蕴薇曹静娟
王健,刘荣堂,张蕴薇,曹静娟
(1.甘肃农业大学草业学院,甘肃 兰州 730070;2.中国农业大学草地研究所,北京 100094)
卫星搭载的植物可产生形态学、物候期、细胞学、生理生化和分子水平等一系列的变异[1]。植物材料在特殊的空间条件下,经受强辐射、微重力、高真空等太空诱变因子的复合作用,发生遗传性状变异[2]。如小麦(Triticum aestivum)出现超绿突变体植株[3],黄瓜(Cucumis sativus)子叶有不同展开度的变异[4],草地早熟禾(Poa pratensis)平均叶片数增加[5],狗牙根(Cynodon dactylon)植株的节间长度变长[6]。除此以外,对空间搭载植物进一步的研究还发现,龙牧803苜蓿(Medicago sativa)M2代的叶绿素含量明显减少[7]。李社荣等[8]研究发现空间搭载的玉米(Zea mays)后代叶片细胞壁薄厚凹凸不平,细胞大小不等,表面不规则,部分细胞退化消失,仅残留细胞壁,细胞液泡化程度加剧,将细胞器挤至四周,并导致叶绿体变形等。本研究以草坪用白三叶(Tri folium repens)为对象,分析卫星搭载对白三叶M1代叶绿素含量、光合特性以及细胞超显微结构的影响,旨为草坪草空间育种提供理论依据。
1 材料与方法
1.1 供试材料供试材料为瑞文德白三叶。精选后的种子分为2份:一份作地面对照(CK),另一份用于卫星搭载(SP)。搭载的“实践八号”育种卫星,于2006年9月9日在酒泉卫星发射中心升空,卫星在近地点187 km、远地点 463 km的近地轨道运行,于9月 24日返回地面。另一部分,贮存于地面温湿度相近(20℃左右)的环境中。
2009年3月随机选取部分种子,在温室用育苗盘育苗,5月10日移栽到花盆中。
1.2 叶绿素的测定称取剪碎处理的和对照白三叶新鲜叶片各0.2 g,转入25 mL棕色容量瓶中,再加入80%的丙酮定容至刻度。将其放入暗室中浸提(浸提过程中摇动2~3次)24 h后,把叶绿体色素提取液倒入光径1 cm的比色杯内,以80%丙酮为空白对照,在721型分光光度计上分别以663、646 nm 波长进行比色,以原提取液为空白溶液,重复3次。按公式计算叶绿素a、b含量[9]。
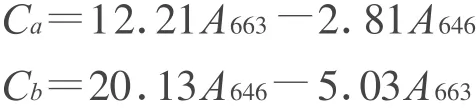

式中,V表示浸提液总体积(mL),W表示样品质量(g)。
1.3 光合特性测定2008年10月1日(晴天)采用便携式光合作用分析仪(LI-6400,美国CID公司)在开路系统下测定各植株瞬时净光合速率(Pn)、气孔导度(Cond)、蒸腾速率(Tr)以及胞间CO2浓度(Ci)。对照和处理各选择5个植株,测定向阳面成熟叶片,分4个时间段:9:00-11:00,11:00-13:00,13:00-15:00,15:00-17:00。计算水分利用效率(WUE=Pn/Tr)[10]。
1.4 叶面积测定在测定光合特性之前,随机选取处理和对照各5株,每一株随机选取3个叶柄上的叶片,用扫描式活体叶面积仪(SHY-150,哈尔滨光学仪器厂)测定,并标记测定的叶片。
1.5 超微结构分别剪取处理和对照相同部位的叶片,切成1 mm2小块,置于用0.1 mol/L磷酸缓冲液(pH值7.4)配制的2.5%戊二醛溶液中进行前固定,用1%锇酸后固定。经0.1 mol/L的磷酸缓冲液冲洗 30%、50%、70%、80%和90%丙酮系列脱水后,用环氧树脂SPURR包埋、聚合。采用切片机(LKB-V,德国徕卡公司)超薄切片,染色,再用(JEM-1230,日本JEOL公司)透射电镜,80 kV加速电压下观察并照相[11]。
1.6 数据分析试验数据处理和对照图形绘制采用Microsoft Excel软件进行,数据的统计分析采用SPSS16.0软件进行。
2 结果与分析
2.1 卫星搭载对白三叶叶片叶绿素的影响
卫星搭载处理对叶绿素a、叶绿素总量有影响,表现出负向变异,即地面对照高于搭载组,卫星搭载白三叶当代植株叶片的叶绿素a和叶绿素总量显著低于对照(P<0.05)。而对于叶绿素b,卫星搭载处理和对照二者间差异不显著(P>0.05);卫星搭载处理叶绿素a、叶绿素b、叶绿素总量的变异程度始终大于对照。
2.2 卫星搭载对白三叶植株光合特性的影响在早上9:00-11:00,处理的胞间CO2浓度差异显著低于对照(P<0.05),在13:00-15:00,净光合速率、气孔导度、蒸腾速率均显著低于对照(P<0.05)。水分利用率(WUE)对照与搭载组间在各个时间段水平均差异不显著(P>0.05)(图1-5)。

图1 卫星搭载对白三叶净光合速率日变化的影响
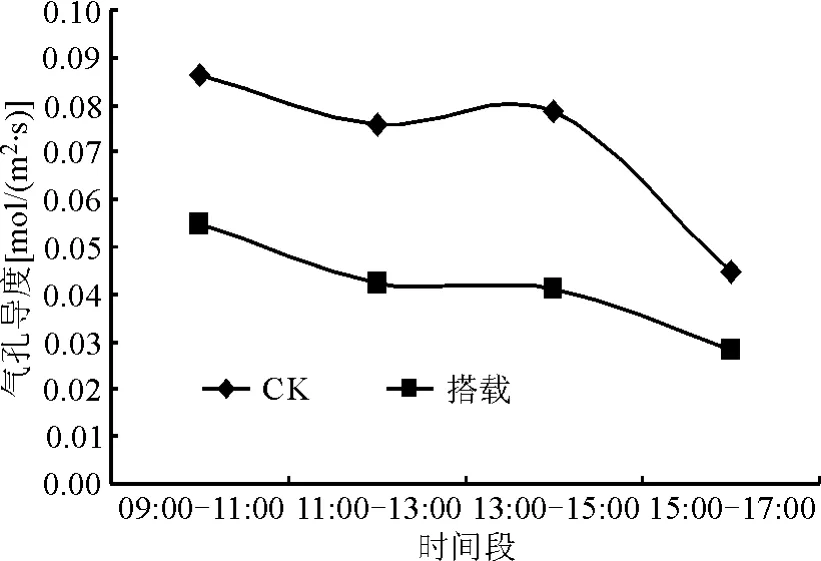
图2 卫星搭载对白三叶气孔导度日变化的影响

图3 卫星搭载对白三叶蒸腾速率日变化的影响

图4 卫星搭载对白三叶胞间CO2浓度日变化的影响

图5 卫星搭载对水分利用率日变化的影响
处理和对照的净光合速率在9:00-11:00最高,而后一直呈下降趋势。但在11:00-13:00、13:00-15:00,下降趋势出现延缓现象。而处理和对照的气孔导度在9:00-11:00最大,对照的蒸腾速率在13:00-15:00最大,在11:00-13:00气孔导度和蒸腾速率同步下降,但 13:00-15:00,气孔导度和蒸腾速率又略微上升。处理和对照的胞间CO2浓度在9:00-11:00最低,而后迅速上升,在13:00-15:00间达到最高。处理和对照的水分利用率与胞间 CO2浓度相反,在9:00-11:00最高,然后也是迅速下降,到下午13:00-15:00下降到最低点,然后开始上升。从图1-3可以明显看到,在1天中的4个时间段,净光合速率、气孔导度和蒸腾速率,处理均比对照低。
2.3 卫星搭载对白三叶植株的超微结构的影响对照与搭载组的白三叶叶片细胞的超微结构差异比较明显,对照白三叶的细胞壁比较均匀,叶绿体也比较均匀的分布在整个细胞中(图6a),淀粉颗粒大,而且比较饱满,较均匀的分布在各个叶绿体中,数量比较少(图6c)。线粒体比较完整(图6e)。基粒片层排列整齐,且贯穿整个叶绿体(图6g)。嗜锇颗粒比较少,且大多数集结在叶绿体中央(图6i)。
卫星搭载组白三叶叶片的细胞,有的细胞壁呈不规则的增厚,扭曲,叶绿体大小不等,有的向细胞中央集中,叶绿体膜有的被撕裂(图6b)。淀粉粒多,且大小不一,有的无序堆积在整个叶绿体中(图6d)。线粒体相对完整,但有的线粒体有明显的破裂,内含物流出,出现空洞、溢裂的现象(图6f)。基粒片层的层数减少,直径却很大(图6h)。推测与叶绿体的性状扭曲有关。嗜锇颗粒在叶绿体中有很大的不同,有的嗜锇颗粒只分布在叶绿体的两侧,且数量极少(图6j),而大多数叶绿体中的嗜锇颗粒比较多,而且比较平均的分布在整个叶绿体中(图6k)。
3 讨论与结论
3.1 卫星搭载对白三叶叶绿素的影响航天诱变能对白三叶光合特性及叶绿素的含量产生影响。有研究[12]发现,草地早熟禾经空间诱变后,3个变异株系叶片光合特性及叶绿素的含量及叶绿素a/b均发生改变,从而影响其对光能的利用效率、固定CO2的能力和对不同光质的利用效率。空间条件引起叶绿素的损伤,或者负向变异较多。主要原因可能是处理引起叶绿素相关酶受到抑制,以及中间产物合成的减少;或者处理引起叶绿素酶作用增强,导致叶绿素的分解加快[13-14]。本研究发现,处理叶绿素总量相对于对照显著下降,其中主要是叶绿素a下降显著。处理叶绿素b相对于对照含量下降,但差异不显著。叶绿素a产生的荧光远大于叶绿素b,而这与光能的吸收,转化能力密切相关[15]。因此,处理一定程度影响了白三叶的光合作用。

图6 卫星搭载对白三叶植株的超微结构的影响
3.2 卫星搭载对白三叶光合特性的影响航天诱变能对白三叶光合特性及叶绿素的含量产生影响。假俭草(Eremochloa ophiuroides)和细叶结缕草(Zoysia japonica)的光合日变化曲线呈双峰型,有“午休”发生[16]。而 Hodges[17]通过研究也发现,由于中午日光辐射强烈,环境相对湿度和CO2含量降低,导致植物光合作用受抑制,净光合速率的日变化曲线会有一个明显的光合午休现象;韩建秋[18]研究发现,土壤相对含水量为25%时,白三叶净光合速率和蒸腾速率下降明显,日变化曲线表现为单峰特征,气孔导度显著降低,而胞间CO2浓度升高,非气孔限制成为Pn下降的主要原因。这与韩烈保等[19]的研究结果一致。本研究中,13:00-15:00,处理的净光合速率、气孔导度、蒸腾速率显著低于对照,处理和对照的气孔导度和蒸腾速率都不呈明显的双峰型,净光合速率处理和对照则一直都呈单峰下降的趋势。这可能是由于在温室中培养,温室的温度过高,引起白三叶土壤水分蒸发过快,从而引起了“干旱胁迫”,造成净光合速率下降。许大全[20]认为,引起叶片光合速率降低的植物自身因素无外乎气孔关闭和叶肉细胞光合活性的下降两类,前者使胞间CO2浓度降低,后者使胞间CO2浓度增高。当2种因素同时存在时,胞间CO2浓度变化的方向依赖于占优势的那个因素。也就是说,当气孔的部分关闭是叶片光合速率降低的主要原因时,胞间CO2浓度降低;而当叶肉细胞光合活性降低是叶片光合速率降低的主要原因时,胞间CO2浓度增高。因此,考虑到上述可能的复杂情况,判断哪一个因素占优势,标准应当是胞间CO2浓度变化的方向。本研究发现,处理和对照净光合速率一直在下降,而处理和对照的胞间CO2浓度则不断上升,说明处理和对照的叶肉细胞光合活性下降,而叶肉细胞光合活性的下降促使净光合速率的下降。在整体上,处理比对照的下降明显,尤其在13:00-15:00,处理的净光合速率显著下降。这可能与处理白三叶的叶绿素a和叶绿素总量显著下降有关。
3.3 卫星搭载对白三叶超微结构的影响空间诱变的红豆草(Onobrychis viciaef olia)叶片的细胞壁不规则的加厚,细胞质稀薄,液泡大,叶绿体变小,形状多为不规则,叶绿体内的淀粉颗粒细小,但数量明显多于对照,匍匐型突变体表现尤为明显[21]。卫星搭载后苜蓿细胞超微结构出现不同程度的变化,搭载组细胞叶绿体扭曲,向细胞中央集中,膜膨散,基粒数目增多,基粒直径增加,片层数减少,叶绿体内嗜锇颗粒数目增加,体积变大,线粒体内外膜膨大或破裂,内含物流出,嵴消失,出现空洞、溢裂现象,叶绿体内淀粉粒数量增多,体积变大,海绵细胞细胞壁明显加厚;个别海绵细胞,淀粉粒几乎占据了整个细胞,数目多,体积大[22]。本研究中,细胞内部的细胞器大多为负向变异,或遭受一定程度的损伤。姚允聪等[23]发现弱光条件下桃(Prunus persica)叶片叶绿体的基粒数与基粒片层数的增加,可能使附着在基粒片层上的补光色素和光合酶增多,有利于光能在类囊体上的传播。但是本研究发现光合效率下降,原因很可能是卫星搭载白三叶叶片细胞中的叶绿体粒片层减少,从而使得嗜锇颗粒在叶绿体中分布不集中,这可能造成了补光色素和光合酶的减少,从而不利于光能的传递。
[1]胡化广,刘建秀,郭海林.我国植物空间诱变育种及其在草类植物育种中的应用[J].草业学报,2006,15(1):15-21.
[2]陈积山,张月学,唐凤兰.我国草类植物空间诱变育种研究[J].草业科学,2009,26(9):173-177.
[3]虞秋成,黄宝才,严建民.作物空间诱变育种的现状及展望[J].江苏农业科学,2001(4):4-6.
[4]李社荣,曾孟浅,刘雅楠,等.植物空间诱变研究进展[J].核农学报,1998,12(6):375-379.
[5]韩曹,孙振元,钱永强,等.神州三号飞船对草地早熟禾生物学特性的影响[J].草业科学,2004,21(4):17-19.
[6]王文恩,包满珠,张俊卫,等.狗牙根辐射诱变后代变异植株的形态特征比较和ISSR分析[J].草业科学,2009,26(12):139-145.
[7]尚晨,韩贵清,陈积山,等.高能混合粒子场处理种子对龙牧 803苜蓿 M2代的抗寒性影响[J].草业学报,2009,18(6):164-168.
[8]李社荣,刘雅楠,刘敏,等.玉米空间诱变效应及其应用的研究Ⅰ.空间条件对玉米叶片超微结构的影响[J].核农学报,1998,12(5):19-25.
[9]朱建玲,徐志防,曹洪麟,等.Cd对南美蟛蜞菊光合特性的影响[J].生态环境,2008,17(2):657-660.
[10]黄顶,张英俊,韩建国,等.白三叶、多年生黑麦草和鸭茅光合生理生态特性研究[J].中国农业大学学报,2006,11(2):44-48.
[11]任丽花,黄敏敏,江枝和,等.干旱胁迫对圆叶决明叶片超微结构的影响[J].中国草地学报,2008,30(1):30-34.
[12]韩曹,孙振元,巨关升,等.空间环境对草地早熟禾诱变效应研究Ⅱ——光合特性和叶绿素含量[J].核农学报,2005,19(6):413-416.
[13]Romero-Puertas M C,Rodrigues-Serrano M,Corpas F J,et al.Cadmium-induced subcellular accumulation of O2·-and H2O2in pea leaves[J].Plant Cell and Environment,2004,27(9):1122-1134.
[14]Lin R Z,Wang X R,Luo Y,et al.Effects of soil cadmium on growth,oxidative stress and antioxidant system in wheat seedlings(Triticum aestivumL.)[J].Chemosphere,2007,69(1):89-98.
[15]孙建云,沈振国.镉胁迫对不同甘蓝基因型光合特性和养分吸收的影响[J].应用生态学报,2007,18(11):2605-2610.
[16]丁小球,胡玉佳,王榕楷.三种草坪草净光合速率和蒸腾速率的日变化特点研究[J].草业科学,2001,18(4):62-68.
[17]Hodges J D.Pattern of photosynthesis under natural environmental conditions[J].Ecology,1967,48:234-242.
[18]韩建秋.水分胁迫对白三叶光合特性的影响[J].上海应用技术学院学报,2009,2(9):106-109.
[19]韩烈保,胡九林,杨永利,等.白三叶草坪蒸散和光合蒸腾速率日变化研究[J].北京林业大学学报,2006,28:22-25.
[20]许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯.1997,33(4):241-244.
[21]张蕴薇,任卫波,刘敏,等.红豆草空间诱变突变体叶片同工酶及细胞超微结构分析[J].草业学报,2004,12(3):223-226.
[22]冯鹏,李键,张蕴薇,等.不同含水量紫花苜蓿种子空载后当代叶片显微和超微结构研究[J].核农学报,2009,23(4):561-565.
[23]姚允聪,王绍辉,孔云.弱光条件下桃叶片结构及光合特性与叶绿体超微结构变化[J].中国农业科学,2007,40(4):855-863.
