封育年限对吉林省西部草地昆虫群落组成结构的影响1)
2010-03-29赵红蕊孟庆繁高文韬
赵红蕊 孟庆繁 高文韬
(北华大学,吉林,132013)
由于过度放牧,近年来吉林省西部草地整体呈退化趋势,局部地区退化严重,导致草地昆虫群落的多样性发生变化,并影响着草地的演替变化[1]。昆虫是草地生态系统的消费者之一,其采食、传粉、分解植物残体和破碎土壤的作用影响着草地植物群落结构演替和植物多样性[2-5]。2000年起吉林省对西部草场采取“封原育草”措施,植灌种草、林草结合,综合治理荒漠化土地,营造乔灌草结合、功能比较完备的复合生态系统,对西部草地恢复起到了促进作用。本试验以吉林省西部草地昆虫群落为研究对象,调查了草地生态恢复过程中昆虫群落组成结构的变化,分析了不同封育阶段昆虫群落的组成特征和影响因素,旨在为西部草地改良和恢复提供科学依据,同时为草场生态恢复过程中害虫种群的控制和昆虫多样性保护及相关研究提供基础资料。
1 研究地概况
吉林省西部草原位于松嫩草原的中部西侧,是欧亚大陆草原在东部的延伸,地理位置为121°18'~129°11'E、43°59'~46°18'N,包括白城市、松原市所辖县(市、区)及农安县、梨树县、双辽市、长岭县等共13个县(市、区),草地总面积467万hm2。吉林省西部草原植被类型的分布和演替与地形地貌、气候、土壤、水分状况、盐分分布及风沙活动等因素有着密切关系。在相同地貌类型上,由于地形变化和土壤水分和盐分状况不同,植被群落也有差异。主要有羊草群丛(Ass.Ley-mus chinensis),虎尾草—羊草群丛(Ass.Leymus chinensis-Chloris virgata),碱蓬—灰蒿群丛(Ass.Artemisia glauca-Suaeda anethifolia),星星草群丛(Ass.Puccinellia tenuiflora)和芦苇群丛(Ass.Phragmites communis)5种植被类型。封育后退化草地得到了较好的恢复,植被盖度已经由恢复前的30%左右恢复到80%以上[6]。
调查样地位于吉林省长岭县八十八乡二十号村和长岭县八十八乡八十八村封育草地试验区。草地在封育前都是退化的平坦天然草原,“封原育草”措施都是在同样基质的草地上进行的,从当地未封育、自然封育2 a、自然封育4 a、自然封育5 a、人工封育5 a草地中选取5块有代表性且初始状况相似的样地,样地间的间隔约5 km,每块样地面积为(50 m×50 m)。调查时各样地植被状况如下:未封育草地最能表现该地区自然生态状况下退化草地类型。草场受人为干扰明显,退化严重。植物群落由9种植物组成,主要有虎尾草(Chloris virgata)、碱蒿(Artemisia anethifolia)、萎陵菜(Potentillae chinensis)、菊科植物(Asteraceae)和羊草(Leymus chinensis)等。植被盖度仅29.5%,平均高度6.06 cm。自然封育2 a草地的植物群落由12种植物组成,主要有虎尾草、蒲公英(Herba taraxaci)、萎陵菜、羊草和碱蒿等,植被盖度42%,平均高度10.33 cm。自然封育4 a草地的植物群落由7种植物组成,主要有羊草、碱蒿、虎尾草、芦苇(Phragmites communis)等。植被盖度34%,平均高度19 cm。自然封育5 a草地植物群落由13种植物组成,主要有羊草、碱蒿、芦苇和蒲公英等。植被盖度38.7%,平均高度12.42 cm。人工封育5 a草地除了使用围栏对草地实行保育措施以外,还在样地内种植了木本植物柽柳(Tamarix chinensis)。植物群落由12种植物组成,主要有羊草、碱蒿、萝藦(Metaplexis japonica)和天门冬(Asparagus cochin-chinensis)等。植被盖度为46.33%,平均高度可达26.07 cm。
2 研究方法
2.1 昆虫群落调查方法
试验于2006—2007年进行,每年5—9月采用样方框、网捕和诱捕法进行昆虫群落调查,每月1次。
样方框调查:在每个样地按单对角线取样法随机选取15个样点,用自制的1 m×1 m的无底样框捕捉样方内昆虫,毒杀,带回室内鉴定。在样地的另一条对角线上设置15个1 m×1 m的小样方,调查样方内植物种类、高度、相对多度和相对盖度。
网捕调查:在每块样地按“Z”字形路线网捕,每样地采集400网/次,将所有捕到的昆虫放入毒瓶,编号,带回室内分类鉴定。
灯诱调查:所选样地都位于不同封育时段草地的近中心。在所选样地,于19:30—23:30时开灯诱集昆虫,诱集半径为150 m,然后编号记载,带回室内分类鉴定。
巴氏罐诱捕调查:在每块样地内设置18个诱杯,其中9个诱杯的引诱剂为腐肉,另9个诱杯的引诱剂为糖醋混合液。先分别在样地的4个顶角、样地边线的中间和样地的中心设置诱杯,引诱剂为腐肉,诱杯编号;然后在每个装腐肉的诱杯旁边隔2 m处再放置1个诱杯,引诱剂为醋、糖、酒精和水的混合物,质量比为2∶1∶1∶20,每个诱杯内放引诱剂40~60 mL,同样记录好编号,同时记录诱杯周围的植物概况,放置诱杯的时间为3 d。
2.2 数据处理
利用Excel统计调查结果,计算多样性指数[7-9],进行优势种分析。即优势种的确定是个体数占总个体数的10%以上,普通种是个体数占总个体数的1%~10%,少于1%的为稀有种。计算方法如下:①物种的丰富度,采用物种数S。②物种多样性指数采用Shannon-Wiener指数
PilnPi。式中:Pi为第i个个体数占群落总个体的比率。③均匀度指数采用Pielou(1967)均匀度指数E=H'/H'max=H'/lnS。式中:H'为Shannon-wiener多样性指数,S为物种数。优势度指数(Dominance Index)采用Berger-Parker指数D=Nmax/Nt。式中:D为优势度指数,Nmax为优势种的种群数量,Nt为群落全部种类的种群数量。优势集中性指数采用Simp-数,ni为物种i的个体数。
3 结果与分析
3.1 封育时间对吉林省西部草地昆虫群落物种组成的影响
从表1可见,草地昆虫物种丰富度随封育时间呈下降的趋势。但是人工封育5 a草地的物种丰富度明显高于自然封育5 a的草地,这可能与人为引种有关。草地昆虫群落优势类群物种丰富度随封育时间的变化呈现较大的差异。鞘翅目昆虫物种丰富度变化趋势与总体物种丰富度变化趋势最为相似,可见鞘翅目类群在群落特征的塑造上具有重要的作用。
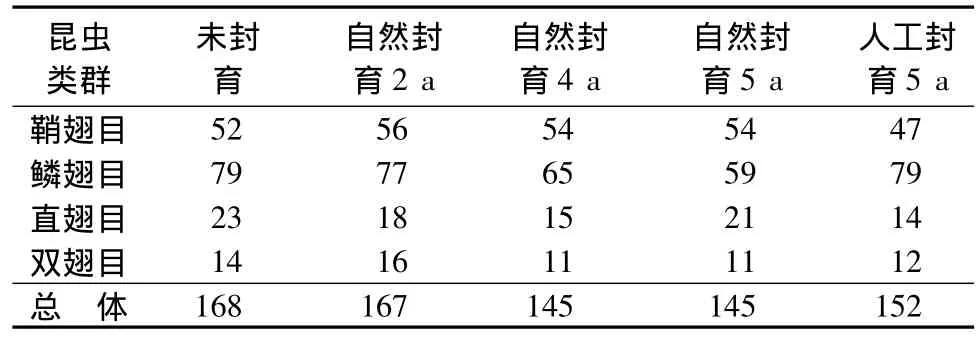
表1 不同封育时段吉林省西部草地昆虫物种组成个
从表2可见,草地昆虫科丰富度随封育时间变化不规则。自然封育2 a的草地昆虫科丰富度较未封育和自然封育4 a和5 a的草地都高,这可能是与调查当年的气候变化有关。人工封育5 a的草地昆虫科丰富度也明显高于自然封育5 a的草地,除了因为气候等环境因子的影响,也可能与人为引种有关。而草地昆虫群落优势类群科丰富度随封育时间的变化呈现较小的差异。直翅目昆虫科丰富度变化趋势与总体科丰富度变化趋势最为相似。

表2 不同封育阶段吉林省西部草地昆虫科组成个
从表3可见,草地昆虫属丰富度随封育时间呈下降趋势。人工封育5 a的草地昆虫属丰富度也比自然封育5 a草地稍高,也可能是人为原因所致。而草地昆虫群落优势类群属丰富度随封育时间的变化呈现较大的差异。鞘翅目昆虫属丰富度变化趋势与总体属丰富度变化趋势最为相似。
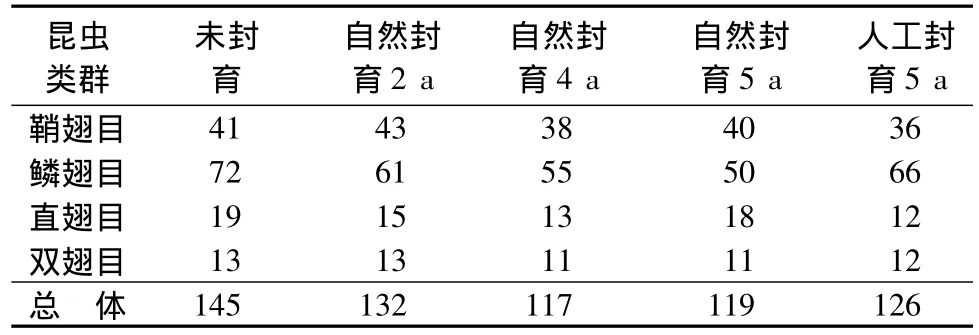
表3 不同封育阶段吉林省西部草地昆虫属组成个
从表4可见,草地昆虫个体数随封育时间出现双峰值。自然封育2 a的草地昆虫个体数高于未封育草地,自然封育4 a草地昆虫个体数最低,自然封育5 a草地昆虫个体数最高且远高于人工封育5 a的草地,这也可能是由于自然和人为原因造成的。而草地昆虫群落优势类群个体数随封育时间的变化呈现较大的差异。鞘翅目昆虫个体数变化趋势与总体个体数变化趋势最为相似。
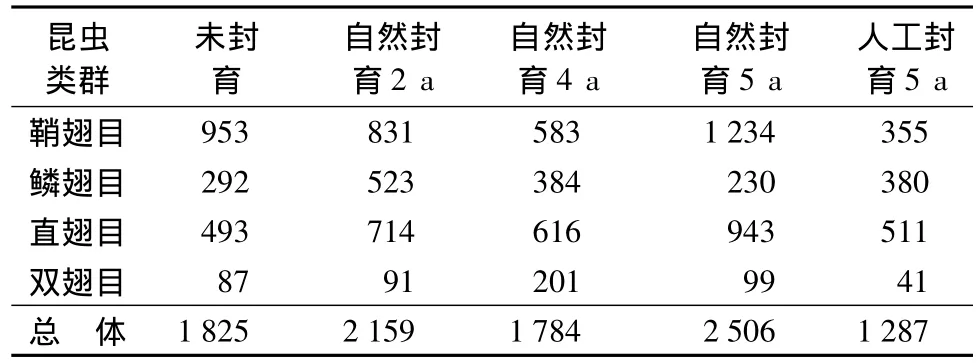
表4 不同封育阶段吉林省西部草地昆虫个体组成个
3.2 不同封育时段草地昆虫群落的结构特征
3.2.1 未封育草地昆虫群落的组成结构特征
从表5可以看出,未封育草地的昆虫个体数的大小顺序为鞘翅目(36.25%)>直翅目(18.75%)>革翅目(17.54%)>鳞翅目(11.11%)>半翅目(5.67%)>膜翅目(4.11%)>双翅目(3.31%)>脉翅目(2.05%)>同翅目(0.49%)=毛翅目(0.49%)>蜻蜓目(0.11%)=螳螂目(0.11%)。而属、种的组成比例则有所不同,鳞翅目>鞘翅目>直翅目>双翅目。优势科为蠼螋科(优势度为0.18)、蝗总科(优势度为0.17)、步甲科(优势度为0.16),三者占科总数的50.29%。该样地共有193种昆虫,寡见种169种,占总数的87.56%,常见种23种,占11.92%,优势种1种,占0.52%。未封育草地生境条件相对严酷,经常有畜群出没,过度放牧较甚,植被破坏严重,盖度较低,而鞘翅目的种群数量比其它类群昆虫占有明显优势,表明鞘翅目昆虫和其它昆虫相比,对环境的适应能力更强。
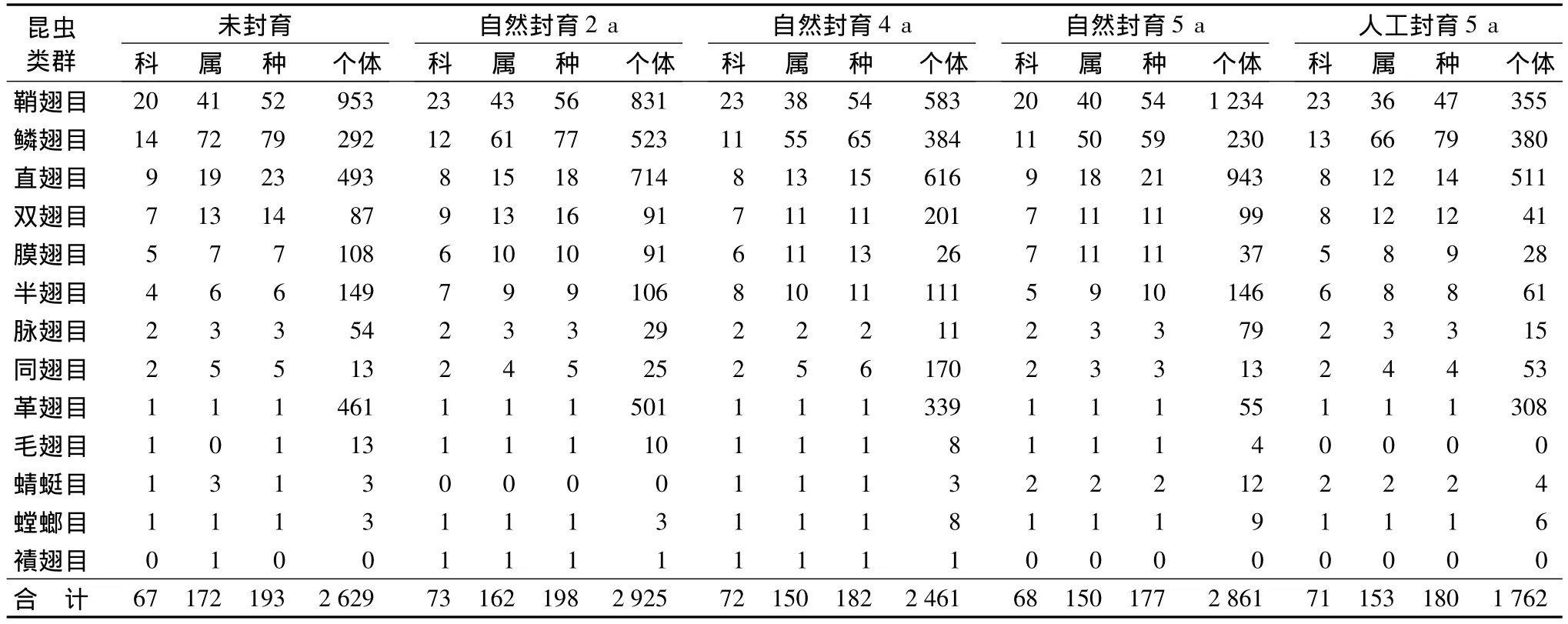
表5 不同封育时段吉林省西部草地的昆虫群落组成个
3.2.2 自然封育2 a草地昆虫群落的组成结构特征
从表5可以看出,封育2 a草地的昆虫个体数的大小顺序为鞘翅目(28.41%)>直翅目(24.41%)>鳞翅目(17.88%)>革翅目(17.13%)>半翅目(3.62%)>双翅目(3.11%)>膜翅目(3.11%)>脉翅目(0.99%)>同翅目(0.85%)>毛翅目(0.34%)>螳螂目(0.10%)>襀翅目(0.03%)。而属、种的组成比例则有所不同,鳞翅目>鞘翅目>直翅目>双翅目。优势科为蠼螋科(优势度为0.17)、斑翅蝗科(优势度为0.15)、步甲科(优势度为0.13)、螟蛾科(优势度为0.11)和夜蛾科(优势度为0.1),占科总数的61.24%。该样地共有物种198种,其中寡见种174种,占个体总数的23.86%,常见种23种,占59.01%,优势种黄褐蠼螋(Labidura sp.),占17.13%。封育2 a的草地,被破坏的植被尚未恢复,生境条件相对比较严酷。鞘翅目和直翅目数量比较接近,占据较大比例,表明这2个目的昆虫有较强的适应性和生存能力。从物种数量看,鳞翅目是最大类群,鞘翅目昆虫步甲占较大类群。步甲是捕食性昆虫,对调节昆虫种群密度有重要意义,在一定程度可以抑制一些害虫种群的爆发,是群落健康稳定发展的重要控制力量。
3.2.3 自然封育4 a草地昆虫群落的组成结构特征
从表5可以看出,封育4 a草地的昆虫个体数的大小顺序为直翅目(25.03%)>鞘翅目(23.69%)>鳞翅目(15.60)>革翅目(13.77)>双翅目(8.17%)>同翅目(6.91%)>半翅目(4.51%)>膜翅目(1.06%)>脉翅目(0.45%)>毛翅目(0.33%)=螳螂目(0.33%)>蜻蜓目(0.12%)>襀翅目(0.04%)。而属、种的组成比例不同,鳞翅目>鞘翅目>直翅目>双翅目。有5个优势科,分别为斑翅蝗科(优势度为0.17)、蠼螋科(优势度为0.14)、螟蛾科(优势度为0.12)、步甲科(优势度为0.10)和叶蝉科(优势度为0.10)。优势种以黄褐蠼螋和大垫尖翅蝗(Epacromius coerulipes(Ivan.))为代表,优势度分别为0.14和0.12,占科总数的25.88%。大垫尖翅蝗啃食植物的叶子,有很强的生存能力和适应性,一旦条件适宜,种群数量常会大爆发,是重要的草原害虫。黄褐蠼螋多为肉食种类,对控制害虫种群爆发有一定的抑制作用。寡见种161种,占总种数的27.71%,常见种19种,占46.40%。封育4 a的草地,植被有了一定程度的恢复,直翅目和鞘翅目数量比较接近,占据较大比例,鳞翅目、革翅目、同翅目、双翅目、半翅目、膜翅目和脉翅目为中等水平,毛翅目、螳螂目、蜻蜓目和襀翅目为小类群,数量较少。从物种数量看,鳞翅目是最大类群,其中主要类群是螟蛾类、夜蛾类和小卷蛾类昆虫。鞘翅目昆虫步甲、虎甲和瓢虫占较大类群,步甲和虎甲大都是捕食性昆虫,以小虫为食,是重要的天敌,一定程度可以抑制一些害虫种群的爆发。
3.2.4 自然封育5 a草地昆虫群落的组成结构特征
从表5可以看出,自然封育5 a草地的昆虫个体数的大小顺序为鞘翅目(43.13%)>直翅目(32.96%)>鳞翅目(8.04%)>半翅目(5.10%)>双翅目(3.46%)>脉翅目(2.76%)>革翅目(1.92%)>膜翅目(1.29%)>同翅目(0.45%)>蜻蜓目(0.42%)>螳螂目(0.31%)>毛翅目(0.14%)。而属、种的组成比例不同,鳞翅目>鞘翅目>直翅目>双翅目。4个优势科为斑翅蝗科(优势度为0.13)、平缘水龟虫科(优势度为0.12)、剑角蝗科(优势度为0.11)和步甲科(优势度为0.11),占科总数的5.88%。常见科有16个,占23.53%;寡见科有48个,占70.59%。常见科数量大小顺序为瓢虫科(4.44%)>槌角蝗科(4.19%)>划蝽科(3.81%)>螟蛾科(3.74%)>虎甲科(3.64%)>夜蛾科(3.39%)>独角仙科(2.66%)>水龟虫科(2.55%)>草蛉科(2.38%)>蚱科(2.20%)>寄蝇科(2.06%)>蠼螋科(1.92%)>葬甲科(1.47%)>鳃金龟科(1.43%)>锥头蝗科(1.12%)。在177个种中有2个优势种,占个体总数的22.47%。其一是平缘水龟虫(Hydrochus sp.),优势度是0.12;另一个是日本鸣蝗(Mongolotettix japonicus(Bolivar)),优势度为0.11。常见种有22种,占个体总数的52.78%;寡见种有153种,占个体总数的24.75%。自然封育5a草地鞘翅目昆虫数量比例最大,是优势类群,其中大部分是步甲、虎甲、瓢虫和水龟虫类。直翅目昆虫所占比例也较大,它的主要类群是鸣蝗类和斑腿蝗类。鳞翅目、半翅目、双翅目、脉翅目和革翅目所占比例比较接近,它们为居间类群;膜翅目、蜻蜓目、同翅目、螳螂目和毛翅目所占比例较小。这里鳞翅目是物种最丰富的类群,它主要是一些螟蛾和夜蛾类。
3.2.5 人工封育5 a草地昆虫群落的组成结构特征
从表5可以看出,人工封育5 a草地的昆虫个体数的大小顺序为直翅目(29.00%)>鳞翅目(21.57%)>鞘翅目(20.15%)>革翅目(17.48%)>半翅目(3.46%)>同翅目(3.01%)>双翅目(2.33%)>膜翅目(1.59%)>脉翅目(0.85%)>螳螂目(0.34%)>蜻蜓目(0.23%)。而属、种的组成比则有所不同,鳞翅目>鞘翅目>直翅目>双翅目。优势科为斑翅蝗科(优势度是0.18)、蠼螋科(优势度是0.17)和夜蛾科(优势度是0.10),占科总数的4.23%。常见科有14个,占科总数的19.18%;寡见科有56科,占76.71%。将常见科按个体数量排序为螟蛾科(9.10%)>步甲科(7.28%)>斑腿蝗科(3.53%)>蟋蟀科(3.30%)>叶蝉科(2.73%)>芫菁科(2.67%)>蚱科(2.16%)>长蝽科(1.88%)>剑角蝗科(1.54%)>尺蛾科(1.42%)>虎甲科(1.25%)>鳃金龟科(1.14%)>花萤科(1.10%)>瓢虫科(1.01%)。优势种分别是黄褐蠼螋(优势度为0.17)和大垫尖翅蝗(优势度为0.11),占个体总数的28.83%。常见种17个,占个体总数的38.20%;寡见种161个,占个体总数的32.97%。人工封育5 a草地以直翅目数量比例最大,特别是大垫尖翅蝗数量较多;鳞翅目、鞘翅目和革翅目数量也明显占优势;半翅目、同翅目、双翅目、膜翅目和脉翅目所占比例比较接近,它们为居间类群,螳螂目和蜻蜓目为小类群,数量较小。鳞翅目是最复杂的类群,其中螟蛾和夜蛾是最主要的类群;鞘翅目中最多的类群是步甲类;双翅目昆虫大部分营腐生生活,草地仍然有放牧现象,牲畜粪便为其提供食物来源,也为其繁育后代提供了温床,故其数量和种类也较多;半翅目中角红长蝽(Lygaeus hanseni Jakovlev)占较大优势;膜翅目中以姬蜂和蚂蚁为主;同翅目的主要类群是叶蝉;脉翅目中主要是草蛉和蚁蛉,蜻蜓目、革翅目和螳螂目物种数较少,不予统计。
3.3 不同封育时段草地昆虫群落物种多样性
在不同封育年限草地类型区,其植被生态环境的差异导致了昆虫群落主要类群所占的比例不同,因此它们在群落中所表现的多样性、均匀性和优势集中性指数也有一定的差异。为进一步分析各草地昆虫群落的变化,对昆虫群落总体特征值进行比较,对各草地昆虫群落生物多样性进行比较分析。
昆虫群落多样性指数变化趋势为人工封育5 a草地(3.8467)>自然封育5a年草地(3.8383)>未封育草地(3.837 8)>自然封育2 a草地(3.810 7)>自然封育4 a草地(3.7896)。均匀度指数变化趋势为自然封育5 a草地(0.741 5)>人工封育5 a草地(0.740 8)>未封育草地(0.729 2)>自然封育4 a草地(0.729)>自然封育2 a草地(0.720 6)。优势集中性指数变化趋势为,人工封育5 a草地(0.056 1)>未封育草地(0.052 3)>自然封育4 a草地(0.051 2)>自然封育2 a草地(0.050 4)>自然封育5 a草地(0.043 8)。可见,人工封育5 a年草地和自然封育5 a草地的昆虫群落多样性指数和均匀性指数均高于其他草地,也就是说这2块草地植被恢复良好,生境比较稳定,适于昆虫群落的生存和繁衍,同时由于种和种之间的相互作用,昆虫群落分布比较均匀。其他草地的昆虫群落多样性指数和均匀性指数并不与封育年限的增加而呈线性相关,是由于吉林省西部草地的土壤条件和低矮植被的差异引起的。从优势集中性指数上看,自然封原育草原对昆虫群落结构产生的较明显影响是,随着封育时间的延长,草地昆虫优势类群较分散,优势度降低。
4 结论与讨论
从不同封育时段的草地昆虫科、属、种和个体数4个水平上分析,自然封育2 a年的草地昆虫明显比未封育草地丰富度指数高,而自然封育4 a的草地昆虫丰富度指数又下降,基本上与未封育草地持平,自然封育5 a的草地昆虫丰富度指数有回升趋势,人工封育5 a的草地昆虫丰富度指数除个体数外都偏高。说明吉林省西部草地的气候、水分和土壤等环境条件和人为干扰特殊,使得封育时段不同,草地昆虫丰富度也不同,为实际生产和物种保护提供重要的基础资料。
从不同封育时段草地昆虫群落组成结构特征来看,自然封育2 a草地虽然生境较差,植被的盖度和高度都比较低,但植物物种数较多。昆虫种群数量较多,其中主要类群是蝗虫、蠼螋、螟蛾和步甲类。自然封育5 a草地是经过5 a的围栏封育,破坏较轻,接近自然状态,植物物种较多,盖度大。其间生存的昆虫种群数量也较高。以鞘翅目、鳞翅目和直翅目数量较多。未封育草地生境最为严酷,这里经常有牧群的出没,过度放牧,植被遭到严重破坏,在一定程度限制了昆虫种群的生存和发展。其中自然封育4 a草地的双翅目昆虫为8.17%,比未封育的数量多,这主要是与其营腐生生活的习性有关。牲畜的粪便为其提供食物来源,一些营寄生和吸血的种类也以牲畜为寄生对象,这些都是双翅目种群获得发展的便利条件。半翅目横纹划蝽数量较多与样地周边的水坝和沼泽有关。自然封育4 a草地植物种类较少,仅有7种植物,该草地的昆虫种群数量比较少。其中主要类群是鞘翅目、革翅目和直翅目。人工封育5 a草地中,人工种植了柽柳,其生存条件更接近于人工环境,受人为影响较大,昆虫类群数量更少,以鳞翅目、直翅目和鞘翅目数量居多。
不同封育时段草地昆虫群落物种多样性差异说明昆虫作为草地植被恢复生态系统中的重要组成部分,其种类、数量与系统内植被群落变化存在着密切的纽带关系。植物群落发生变化,昆虫群落也随之发生适应性变化。观察昆虫优势集中性指数可以看出,人工封育5 a草地受人为干扰影响较大,优势集中性较高,其它草地随着植被恢复和种类的增加,有了一个较大的物种生境和多样化的食物链资源,昆虫优势集中性降低。这主要是由于人工封育5 a草地经过人工群落建造,增加了乔木物种,又有围栏封育的养护,使环境资源的质和量相应改变,扩大了生态容量,创造了优良而稳定的生态环境,从而增大了昆虫栖境的多样性,并使各种昆虫组成了一个较为复杂的群落,其昆虫群落的多样性指数均大于其他类型草地。随着草地封育年限的增加,植被较好,覆盖度高,为许多昆虫提供了良好的栖息场所,昆虫群落多样性增加,而封育2 a草地多样性偏高是由于虽然植被盖度较小,但低矮植物较多,植被物种比较丰富,而植物种类的多少及变化直接影响昆虫群落的营养通道,使得昆虫群落对环境的适应产生波动,多样性也产生波动。并且春季气候转暖,植被刚刚复苏,封育年限高一点的草地植被长势略好,有利于形成比较均匀的昆虫群落。
综上所述,在不同封育时段草地类型中,其植被生态环境的差异导致了昆虫群落主要类群组成、物种多样性和优势集中性指数的差异。草地昆虫物种丰富度指数随封育时间延长呈下降的趋势。这可能与封育时间延长,植被盖度增高,物种组成变化有关。
[1]Parker M,Mac Nally R.Habitat loss and the habitat fragmentation threshold:an experimental evaluation of impacts on richness and total abundances using grassland invertebrates[J].Biol Conserv,2002,105:217-229.
[2]De Deyn G B,Raaijmakers C E,Zoomer H R,et al.Soil invertebrate fauna enhances grassland succession and diversity[J].Nature,2003,422:711-713.
[3]Hemerik L,Brussaard L.Diversity of soil macro-invertebrates in grasslands under restoration succession[J].Eur Soil Biol,2002,38(2):145-150.
[4]Koricheva J,Mulder C P H,Schmid B,et al.Numerical responses of different trophic groups of invertebrates to manipulations of plant diversity in grasslands[J].Oecologia,2000,125:271-282.
[5]Watkinson A R,Ormerod S J.Grasslands,grazing and biodiversity:editors'introduction[J].J Appl Ecol,2001,38:233-237.
[6]刘泉波.吉林省西部退化草地改良恢复研究[D].长春:吉林大学,2005.
[7]张金屯.植被数量生态学方法[M].北京:中国科学技术出版社,1995.
[8]杨玉盛,何宗明,邱仁辉,等.严重退化生态系统不同恢复和重建措施的植物多样性与地力差异研究[J].生态学报,1999,19(4):490-494.
[9]李青,吴兆录,刘玲玲,等.滇西北藏区草地管理方式对草地昆虫群落多样性的影响[J].生态学杂志,2006,25(11):1375-1379.
