真江蓠杭州湾海域栽培试验及生态因子对藻体生长的影响
2010-03-15霍元子徐姗婻张建恒何培民
霍元子,徐姗婻,张建恒,韩 芳,董 丽,何培民
(上海海洋大学水产与生命学院,水域环境生态上海高校工程研究中心,上海 201306)
真江蓠(Gracilaria verrucosa)属于红藻门(Rhodophyta),藻体大,生长快,含胶量高,可达 30%以上,是制造琼胶的重要原料之一[1],除了食用和作为药材原料外,也可用作鲍鱼的饵料[2]。我国北起辽东半岛,南到广东南澳县、西到广西防城港市均有分布[3]。近年来以高浓度N、P为主要特征的海域富营养化已成为全球近海海域普遍存在的环境问题[4]。杭州湾属我国严重污染海域之一,多年来无机氮磷等污染物持续超标[5]。已有的研究证实[6~9],在近海海域栽培大型海藻,是一种对环境进行原位修复的有效手段[10]。目前,有关江蓠属的研究多集中于细基江蓠繁枝变型(G.tenuistipitata)[11~13]、菊花心江蓠(G.lichenoides)[14]和龙须菜(G.lemaneiformis)[15~18]等 3种。在低盐且含沙量高的杭州湾上海海域试栽培真江蓠并对其适宜生长的环境因子进行研究尚属首次。本文研究报道了真江蓠杭州湾海域栽培试验以及温度、盐度和光照强度等生态因子对真江蓠生长影响的实验结果,为大规模栽培及其进一步应用于近海海域富营养化的生物治理提供理论依据。
1 材料与方法
1.1 材料
真江蓠(G.verrucosa)小苗来自上海海洋大学藻类学实验室,于2003年5月采自浙江省宁波奉化象山港湖头渡的网箱养殖区,编号为 YSJ03-01,保种于无菌的培养液中,温度为 20~25 ℃,盐度 20,光强 30~50 µmol/(m2·s),光照周期 12L:12D,每隔 1 周左右更换经加富的培养液1次。
1.2 藻体培养
将株系 YSJ03-01进行大量培养,培养容器为1~5 L三角烧瓶,培养液为自然海水加富VSE,盐度20 ± 0.2,pH 8.0。培养条件为:温度 25 ℃,光强30~50 µmol/(m2·s),光照周期12L:12D,空气经CuSO4过滤后充于培养瓶中。在实验前1周培养,供实验用。
1.3 室内生态因子实验设计
将旺盛生长真江蓠,切成长度约为 10 cm的藻体,放入盛有200 mLVSE培养液的250 mL三角烧瓶中,每瓶中放置真江蓠的初始质量为0.23 g±0.01 g。温度设置 10、15、20、25、30、35 ℃共 6个梯度,盐度设置10、15、20、25、30共5个梯度,光照强度设置 30、60、90、120 µmol/(m2·s)共 4 个梯度,共120个温度-盐度-光照强度组合,每组设置3个重复,空气经CuSO4过滤后充于培养瓶中。实验时间为7 d,隔天加富营养1次。
1.4 真江蓠海区栽培实验
真江蓠的海区栽培实验在杭州湾北岸上海市金山区近岸一围隔海域内进行,面积为1.72 km2。栽培区共分 3块,每块面积为 250 m×60 m,每块间隔60~100 m,每块由21根250 m长缆绳组成,缆绳之间距离为3 m。缆绳两端用竹桩固定(缆撅),中央部分用 5~6个浮子等距离支撑。采自宁波象山养殖区的3.5 t 新鲜真江蓠苗种运至海区后,即按每10 kg平铺式装入孔径为0.5 cm,规格为0.5 m×10 m,经加工缝合的聚乙烯网袋中。网袋口用聚乙烯绳缝合,间隔将网袋长边悬挂在 250 m长的缆绳上,每条缆绳悬挂 5~6个装有真江蓠的网袋,相邻缆绳上的网袋间隔排挂。通过在缆绳上悬挂重物,调整网袋位置为水面以下1~2 m。栽培时间由2006年8月下旬至2007年7月。
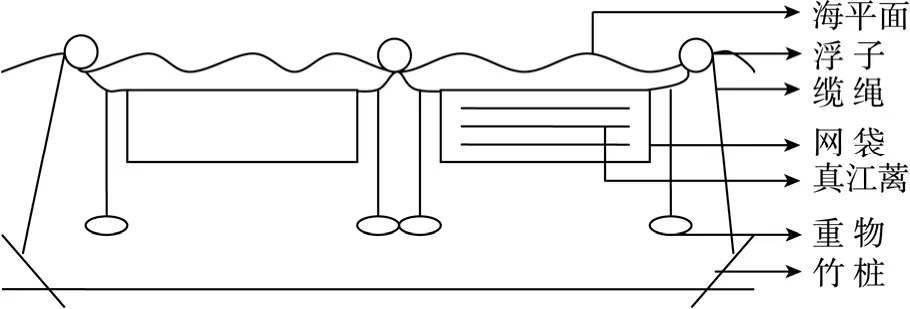
图1 真江蓠网袋养殖栽培模式示意图Fig.1 Scheme of culturing G.verrucosa through mesh bag
1.5 真江蓠生长的测定
1.5.1 室内实验藻体生长的测定
藻体离水后用吸水纸吸干称其湿质量,藻体湿质量用10-4g电子天平称量,按下式计算真江蓠的特定生长率RSG(%/d)[19]和相对生长率RRG(%)[20]:
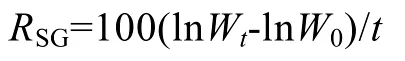
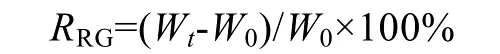
其中,W0为真江蓠初始鲜质量(g),Wt为实验进行至第t天时的真江蓠鲜质量(g)。
1.5.2 海区栽培实验藻体生长的测定
对网袋中的真江蓠定期称质量:栽培前,记录每袋真江蓠的位置和质量; 实验开始后,每 14天从缆绳上随机解下3~5袋真江蓠,沥干水分进行称量。根据前后称量结果,按(1)式计算真江蓠的特定生长率RSG(%/d)。
1.6 数据分析
试验数据用 SPSS 13.0统计分析软件进行方差分析,再采用Duncan多重比较分析试验结果均值的差异显著性(P< 0.05)。
2 结果
2.1 真江蓠生长多生态因子方差分析结果
根据多因素方差分析结果(表 1),温度、盐度和光照强度对真江蓠的生长都有极显著影响(P<0.01)。从均方值来看,不同温度造成的组间差异最大,这说明真江蓠生长对温度的变化最敏感; 其次是盐度,对真江蓠的生长影响也较明显; 相对而言,光照强度对其生长的影响没有前2者强烈。3种生态因子中,任意2因子之间都具有显著交互效应(P<0.05),特别是温度和盐度的交互效应极显著(P<0.01),且3个生态因子之间的交互效应也极显著(P<0.01),说明这 3个生态因子对真江蓠生长的影响不是相互独立的。
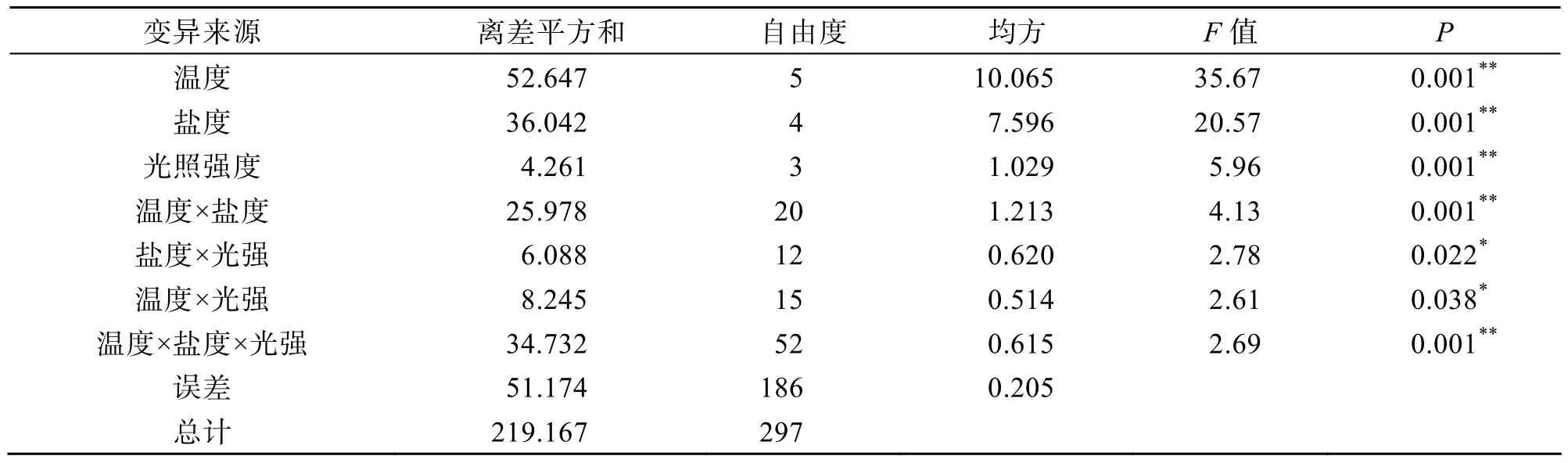
表1 温度、盐度和光照强度对真江蓠生长影响的方差分析Tab.1 Effects of temperature,salinity and light intensity on the growth of G.verrucosa
2.2 真江蓠生长单因子分析
2.2.1 温度对真江蓠生长的影响
温度对真江蓠藻体生长的影响尤其明显(表2)。在 10℃时,真江蓠藻体生长最慢,显著低于其他温度组。在15℃和20℃也存在显著差异,生长速度随温度升高而提高。温度在20~30℃之间时,生长无显著差异,生长率较高(128.93 %~141.45 %),也相对比较平稳,但随着温度的继续上升,温度为 35℃时,真江蓠生长率呈现出下降趋势。由此得知,真江蓠在温度为20~30 ℃范围内生长较好,其中25℃时最好。
2.2.2 盐度对真江蓠生长的影响
盐度对真江蓠生长影响较大(表3)。真江蓠藻体在低盐度10和较高盐度30培养液中时,其生长率均低,与其他盐度组存在显著差异; 盐度为15~25之间时,真江蓠相对生长率明显提高,均超过100 %,其中盐度为 20时最高(128.33 %),后随盐度升高而降低。可见真江蓠藻体生长适宜盐度范围在 15~25之间。
2.2.3 光照强度对真江蓠生长的影响
光照强度对真江蓠生长的影响见表4,光照强度在30 和 60 µmol/(m2·s)之间时,真江蓠生长差异显著,而当光强达到 90 µmol/(m2·s)时,生长率最高(120.97 %),显著大于 30 µmol/(m2·s)和 120 µmol/(m2·s)组的生长情况。因此,适宜真江蓠生长的光照强度应在 90µmol/(m2·s)左右。

表2 不同温度对真江蓠生长的影响Tab.2 Effect of temperature on the relative growth rate of G.verrucosa

表3 不同盐度对真江蓠生长的影响Tab.3 Effect of salinity on the relative growth rate of G.verrucosa

表4 不同光照强度对真江蓠生长的影响Tab.4 Effect of light intensity on the relative growth rate of G.verrucosa
2.3 真江蓠生长多因子交互效应分析
2.3.1 温度和盐度的交互效应
温度和盐度交互作用对真江蓠生长的影响见图2。结果表明,在 10~35℃范围内,盐度为 10时,真江蓠生长率均为最低,而盐度为 20时,真江蓠生长率均为最高,盐度为15和25时,真江蓠生长率均次于盐度20。生长最好组合为温度为25℃和盐度为20。

图2 温度和盐度对真江蓠生长的影响Fig.2 Combined effect of temperature and salinity on the relative growth rate of G.verrucosa
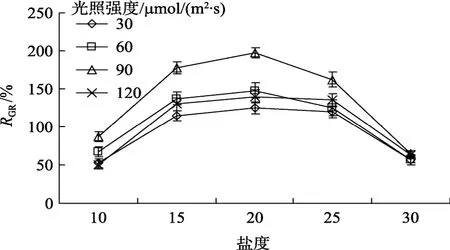
图3 盐度和光强对真江蓠生长的影响Fig.3 Combined effect of salinity and light intensity on the relative growth rate of G.verrucosa
2.3.2 盐度和光强的交互效应
盐度和光照强度交互作用对真江蓠生长的影响见图 3。由图可知,在盐度 10~30,光照强度 30~90µmol/(m2·s)范围内,真江蓠生长率均随光照强度提高而逐步提高,光照强度为 90 µmol/(m2·s)时,生长率最高。最佳盐度和光强组合为盐度20和光照强度90 µmol/(m2·s)。
2.3.3 温度和光强的交互效应
温度和光照强度交互作用对真江蓠生长的影响(图 4)表明,在光照强度 30~90 µmol/(m2·s)和 10~25℃范围内,真江蓠生长率均随温度上升和光强增加而提高,超过该范围后生长速率急速下降,35 ℃时生长最慢。对真江蓠而言,生长的光照强度和温度最佳组合为 90 µmol/(m2·s)和 25 ℃。
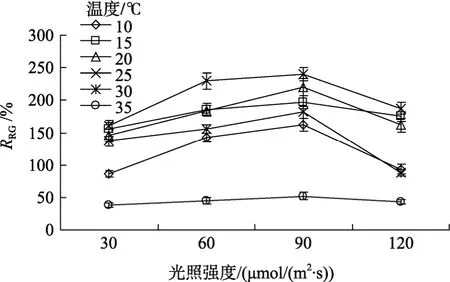
图4 温度和光强对真江蓠生长的影响Fig.4 Combined effect of temperature and light intensity on the relative growth rate of G.verrucosa
2.4 真江蓠在杭州湾海域的生长情况
2006年8月~2007年7月,夏季围隔海区平均水温在 28.93~31.80℃之间,冬季平均水温为 5.20~11.80℃(图 5)。2006 年 9~11 月和 2007 年 4~6 月真江蓠在金山城市沙滩围隔海区的生长率显著高于其他月份(P<0.05),最高生长率出现在9月和5月,RSG分别为15.88 %/d和15.16 %/d。2006年12月底采收了 5.3 t真江蓠(以鲜质量计),仅保留少部分真江蓠在围隔海区栽培筏架上继续栽培,以观察越冬生长情况。1月围隔海区水温最低为0~1 ℃,真江蓠生长曾出现停滞或负增长状况,但随着2月气温上升,栽培越冬的真江蓠生物量增长了近 1倍,并将其作为苗种继续进行扩大面积栽培。实验结果表明真江蓠可以在上海金山围隔海区度夏和越冬。

图5 真江蓠在大型围隔海区的生长率变化Fig.5 Variation of the specific growth rate of G.verrucosa in a large enclosed sea area
3 讨论
上海曾在80年代末和 90年代初从福建引进过江蓠,但栽培未获得成功,本研究为首次在杭州湾上海金山海域成功实现真江蓠的栽培。金山海域位于杭州湾北岸,受钱塘江涌潮影响,平均潮差2.5~3.4 m,最大潮差达7.5 m; 水体泥沙含量高,平均含沙量达到0.5~1.0 kg/m3,2006年3月实测的海水盐度平均为 10。一般江蓠栽培通常选择水色澄清、透明度较大的海区以保证适宜的光照强度。本研究最初在杭州湾上海金山海域进行真江蓠移栽时,曾采用粗绳夹苗的栽培方式,但由于受到强光照射和敌害生物附生等因素影响,首批真江蓠苗断落或生长不佳,死亡率大于 80 %; 设计并实施全浮动式网袋栽培后,可有效防止上述生长不利因素。真江蓠在金山围隔海区全年平均特定生长率(RSG)为9.42 %/d。生长率受水温影响显著,当水温在 16~29 ℃时,生长最快,RSG最高出现在 9月和 5月,分别为15.88 %/d 和 15.16 %/d; 而低温(<15 ℃)和高温(>30℃)下其生长率均显著下降; 受到冬季 1月围隔海区较低水温的(最低为 0~1 ℃)影响,真江蓠生长虽曾出现停滞或负增长状况,但仍可在气温上升后继续生长和作为苗种进行扩大面积栽培。真江蓠全年平均生长率略低于作者 2006年夏季在象山港网箱区养殖的真江蓠生长率(平均RSG为 11.52 %/d)[21],以及 Zhou等[22],Yang等[16]报道龙须菜的生长率(11~13 %/d),但高于Troell等[23]在鲑鱼网箱养殖区的结果(7 %/d)。本实验结果表明,真江蓠在杭州湾上海海域已能正常生长。
江蓠属于广温性种类[16]。本实验结果表明,在10~35 ℃真江蓠均能存活和生长,在15~25 ℃的范围内随温度升高真江蓠生长率增加,大于30 ℃以后生长率下降,这表明其最适生长温度在 25 ℃左右,过高或过低的温度都不利于真江蓠的生长。该结果与刘静雯等[12]、刘思俭[3]等对细基江蓠繁枝变型生长适宜温度范围的研究结果较为接近。但在研究温度、盐度和光照强度 3因子交互效应时,真江蓠 30℃时平均相对生长率仍高达 128.93 %,藻体无明显异常现象; 而 35 ℃时藻体颜色加深且生长趋缓,平均相对生长率亦达84.60 %,表明真江蓠在短期高温下生长减缓,但仍能存活,有利于种苗在栽培海区安全渡夏。
江蓠也属广盐性种类[2],喜生长在有淡水流入和水质肥沃的内湾中。本实验中真江蓠能够在盐度为10下存活和生长,盐度为15~25为真江蓠适宜生长的范围,说明真江蓠耐低盐性较好。因此在河口附近海域,由于海水盐度较低,真江蓠能够大量旺盛生长。比细基江蓠繁枝变型的耐盐范围更广,但在纯淡水中和该种一样,20天后即全部死亡[2]。
江蓠喜生长于光线充足的潮间带石沼中,主要营固着生活,可固定在岩石和石砾等各种基质上,不能主动通过垂直移动来适应光强变化,因此光强也是决定其分布区域的主要因素之一[1]。光照强度对真江蓠生长的影响没有温度和盐度大,本实验表明真江蓠在光强 30~90 µmol/(m2·s)之间,随光强增加,生长呈增快趋势。
总体来看,温度、盐度和光照强度对真江蓠的生长影响显著,且具有显著的交互作用。本实验结果得出,真江蓠生长最适条件为温度25 ℃,盐度20和光强为90 µmol/(m2·s)的组合。真江蓠对温度、盐度和光强等生态因子的耐受范围比较广,具有较强的生存能力,可作为候选的生态修复藻种施用于近岸海域的富营养化治理。但除了温度、盐度和光照强度外,其他生态因子,如营养盐[24,25]、捕食者[26],对江蓠属大型海藻的生长亦有显著的影响。因此,关于这些生态因子对真江蓠生长的影响还有待于进一步研究。
[1]曾呈奎,王素娟,刘思俭,等.海藻栽培学[M].上海:科技出版社,1985.225-250.
[2]刘思俭.我国江蓠的种类和人工栽培[J].湛江海洋大学学报,2001,21(3):71-79.
[3]刘思俭.江蓠养殖[M].北京:农业出版社,1988.36-68.
[4]Nixon S W,Ammerman J W,Atkinson L P,et al.The fate of nitrogen and phosphorus at the land-sea margin of the North Atlantic Ocean [J].Biogeochemistry,1996,35:141-180.
[5]国家海洋局.2000~2009年中国海洋环境质量公报.http://www.soa.gov.cn/hyjww/hygb/hyhjzlgb/A020707 index_1.htm,2010-06-10.
[6]Chopin T,Yarish C,Wilkes R,et al.DevelopingPorphyra/salmon integrated aquaculture for bioremediation and diversification of the aquaculture industry [J].Journal of Applied Phycology,1999,11:463-472.
[7]Chapman A R O,Craigie J S.Seasonal growth inLaminaria longicruris:Relations with dissolved inorganic nutrients and internal reserves of nitrogen [J].Marine Biology,1997,40:197-205.
[8]Neori A,Cohen I,Gordin H.Ulva lactucabiofilters for marine fishpond effluents:II.Growth rate,yield and C:N ratio [J].Botanica Marina,1991,34:483-489.
[9]Martinez-Aragon J F,Hernandez I,Perez-Llorens J L,et al.Biofiltering efficiency in removal of dissolved nutrients by three species of estuarine macroalgae cultivated with sea bass (Dicentrarchus labrax) waste waters I.Phosphate [J].Journal of Applied Phycology,2002,14(5):365-374.
[10]杨宇峰,宋金明,林小涛,等.大型海藻栽培及其在近海环境的生态作用[J].海洋环境科学,2005,24(2):78-80.
[11]吴超元,李纫芷,林光恒,等.细基江蓠繁枝变型生长适宜环境条件的研究[J].海洋与湖沼,1994,25(1):60-66.
[12]刘静雯,董双林.光照和温度对细基江蓠繁枝变型的生长及生化组成影响[J].青岛海洋大学学报,2001,31(3):332-338.
[13]刘静雯,董双林,马.温度和盐度对几种大型海藻生长率和 NH4-N吸收的影响[J].海洋学报,2001,23(2):109-116.
[14]牛化欣,马,田相利,等.菊花心江蓠对中国明对虾养殖环境净化作用的研究[J].中国海洋大学学报,2006,36(Sup.):45-48.
[15]Fei X G.Solving the coastal eutrophication problem by large scale seaweed cultivation [J].Hydrobiologia,2004,512:145-151.
[16]Yang Y F,Fei X G,Song J M,et al.Growth ofGracilaria lemaneiformisunder different cultivation conditions and its effects on nutrient removal in Chinese coastal waters[J].Aquaculture,2006,254:248-255.
[17]汤坤贤,游秀萍,林亚森,等.龙须菜对富营养化海水的生物修复[J].生态学报,2005,25(11):3044-3051.
[18]汤坤贤.龙须菜在网箱养殖区的生物修复研究[J].中国水产科学,2007,14(3):488-492.
[19]Metaxa E,Deviller G,Pagand P,et al.High rate algal pond treatment for water reuse in a marine fish recirculation system:Water purification and fish health [J].Aquaculture,2006,252:92-101.
[20]张寒野,吴望星,宋丽珍,等.条浒苔海区试栽培及外界因子对藻体生长的影响[J].中国水产科学,2006,13(5):781-786.
[21]徐姗楠,温珊珊,吴望星,何培民.真江蓠(Gracilaria verrucosa)对网箱养殖海区的生态修复及生态养殖匹配模式[J].生态学报,2008,28(4):1466-1475.
[22]Zhou Y,Yang H S,Hu H Y,et al.Bioremediation potential of the macroalgaGracilaria lemaneiformis(Rhodophyta) integrated into fed fish culture in coastal waters of north China [J].Aquaculture,2006,252:264-276.
[23]Troell M,Halling C,Nilsson A,et al.Integrated marine cultivation ofGracilaria chilensis(Gracilariales,Rhodophyta) and salmon cages for reduced environmental impact and increased economic output[J].Aquaculture,1997,156:45-61.
[24]林贞贤,宫相忠,李大鹏.光照和营养盐胁迫对龙须菜生长及生化组成的影响[J].海洋科学,2007,31(11):22-26.
[25]黄鹤忠,孙菊燕,申 华,等.无机氮浓度及其配比对细基江蓠繁枝变型生长及生化组成的影响[J].海洋科学,2006,30(9):23-27.
[26]郑新庆,姚雪芬,黄凌风,等.厦门 湖龙须菜生长的主要影响因素[J].海洋科学,2008,32(12):47-51.
