猪乳铁蛋白基因的克隆及其重组乳酸菌表达系统构建
2010-02-20唐丽杰赵丽丽宗晓淋刘荻乔薪媛姜艳萍葛俊伟李一经单安山
唐丽杰,哈 卓,赵丽丽,宗晓淋,刘荻,乔薪媛,姜艳萍,葛俊伟,李一经,单安山
(1.东北农业大学生命科学学院,哈尔滨 150030;2.东北农业大学动物医学学院,哈尔滨 150030;3.东北农业大学动物科学技术学院,哈尔滨 150030)
乳铁蛋白(Lactoferrin,LF)是一种天然活性铁结合糖蛋白,广泛存在于动物的乳汁(初乳)、血液、唾液、小肠液等外分泌液或血浆、中性粒细胞中。其中除乳腺组织中含量丰富外,其他组织中均含量甚微。LF是一种具有多种生物学功能的蛋白质,它不仅参与铁的转运,而且具有抗微生物、抗氧化、调节免疫系统等功能。在动物生产中使用乳铁蛋白及其酶解产物乳铁蛋白活性肽(Lactoferricin,LFcin),可以降低仔猪的缺铁性贫血发病率、提高断乳仔猪的免疫机能和改善肠道微生态环境,促进断乳仔猪健康生长。
鉴于乳铁蛋白的广泛生物学活性,目前在高效补铁剂、饲料添加剂、免疫调节剂以及抗氧化剂等方面正广泛应用,但是由于传统的乳铁蛋白分离从初乳中进行提取纯化工艺繁琐,因此以基因工程手段生产乳铁蛋白是该技术的主要方法[1-6]。作为基因工程的宿主菌无论是大肠杆菌、酵母菌,还是动物细胞,表达的外源蛋白大多需纯化,而繁琐的纯化工艺一直是基因工程产品不能应用于生产实践的障碍。
而乳铁蛋白对于铁需求低的乳酸菌基本不抑制,且能促进动物肠道中双歧杆菌和乳酸杆菌等有益菌群生长;作为载体传递系统的干酪乳杆菌是一种益生菌,国内外许多研究结果表明其可作为一种理想的表达细胞,具有目前所知的其他真核或原核生物无法媲美的优越性。因此本研究以泌乳猪乳腺组织RNA为模板,获得猪乳铁蛋白基因的克隆,同时以乳杆菌质粒作为表达载体,构建表达猪乳铁蛋白基因的多种重组乳酸菌,为获得猪乳铁蛋白的乳酸菌表达及筛选最佳生物学活性的重组猪乳铁蛋白奠定基础。
1 材料与方法
1.1 材料
大肠杆菌和乳酸菌穿梭表达分泌型载体质粒pPG612.1,Lactobacillus casei ATCC393由荷兰Jos Seegers博士惠赠;Lactobacillus plantarum KLDS 1.0344,Lactobacillus paracasei KLDS1.0 652,Lactobacillus pentosus KLDS1.0413源自乳品科学教育部重点实验室;pMD18-T载体购自大连宝生物工程有限公司;大肠杆菌JM109感受态细胞由本实验室保存。
1.2 引物
应用Oligo6.0软件设计引物,引物PF1、PF2用于RT-PCR从乳腺组织中扩增猪乳铁蛋白基因N端序列,并分别含有Bam HⅠ和XholⅠ酶切位点。引物由上海生物工程有限公司合成,引物序列如下:PF1:5′GAGCTCAATGAAGCTCTTCATCCC 3′;PF2:5′CTCGAGCTTCGCCTGCCGCGC 3′。
1.3 猪乳腺组织RNA的提取
取泌乳3 d的长白猪乳腺组织在液氮中研磨成粉末,按照每50~100 mg组织加入1 mL Trizol液裂解乳腺组织细胞,进一步使用氯仿抽提及异丙醇沉淀提取RNA,-20℃备用。
1.4 RT-PCR扩增猪乳铁蛋白基因N端序列
利用设计合成的P2引物进行反转录,获得cDNA,并以其为模板,按照如下过程进行PCR扩增猪乳铁蛋白基因N端序列,95℃预变性5 min,94℃1 min,57℃1 min,72℃1 min,35个循环,72℃延伸10 min;反应结束后用1.0%琼脂糖凝胶电泳检查扩增结果。将获得的基因命名为PLFN。
1.5 PLFN基因的克隆
将获得的PLFN基因PCR产物经胶回收纯化后与pMD18-T载体进行16℃连接过夜;连接产物将用于转化JM109感受态细胞。在LB琼脂平板上挑取菌落接种于 5 mL 含 100 μg·mL-1Amp的LB 液体培养基中,37℃震荡培养过夜,按《分子克隆实验指南》的碱裂解法小剂量制备质粒的步骤提取质粒。对重组质粒进行Sac I单酶切,Sac I、Xhol I双酶切鉴定,同时进行PCR鉴定及序列测定分析。阳性重组质粒命名为 pMD18-T-PLFN。
1.6 带有粘性末端的目的基因片段及干酪乳杆菌表达载体的制备
用小量质粒DNA提取试剂盒(上海华瞬生物工程有限公司)从大肠杆菌JM109提取重组质粒pMD18-T-PLFN。从干酪乳杆菌Lactobacillus casei 393中提取质粒pPG612.1,按Anderson等方法从乳酸菌中提取质粒[7-8]。将干酪乳杆菌表达载体pPG612.1和重组质粒pMD18-T-PLFN分别进行SacⅠ、XholⅠ双酶切,结果见表1。
置于37℃水浴反应4 h。1.0%琼脂糖凝胶电泳分析酶切结果。然后分别对目的片段用柱式胶回收试剂盒(上海华瞬生物工程有限公司)分别回收3 500 bp的载体片段和1 000 bp的基因片段。
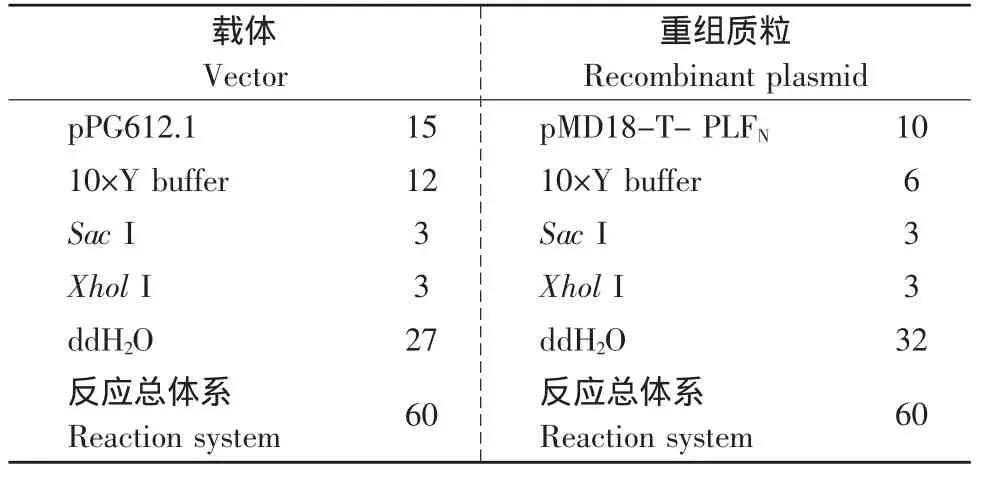
表1 酶切体系Table 1 Reaction system of restriction enzyme(μL)
1.7 PLFN基因干酪乳杆菌表达载体的构建
1.7.1 双酶切纯化产物的连接
将1.6制备的双酶切纯化产物进行连接。连接反应体系为粘性末端pPG612.1 5 μL,10×T4Ligase buffer 2 μL,T4DNA Ligase 1 μL,ddH2O 2 μL,粘性末端S基因10 μL,将连接反应体系混匀。置于连接仪中16℃连接过夜;连接产物将用于转化JM109感受态细胞。在LB琼脂平板上挑取菌落接种于 5 mL 含 10 μg·mL-1氯霉素(Cm)的LB 液体培养基中,37℃震荡培养过夜,按《分子克隆实验指南》的碱裂解法小剂量制备质粒的步骤提取质粒。对重组质粒进行Sac I单酶切,Sac I、Xhol I双酶切鉴定,同时进行PCR鉴定。将重组质粒命名为pPG612- PLFN。
1.7.2 电转化用干酪乳杆菌感受态细胞的制备及电转化
乳酸杆菌ATCC393、KLDS1.0677、KLDS1.0652及KLDS1.0413按照如下方法制备感受态细胞:挑取新鲜培养的单菌落,接种于5 mL MRS液体养基中,37℃过夜静止培养;取过夜培养物5 mL,用MRS培养液稀释至200 mL,37℃继续培养至OD590为0.6左右;将培养液转移到预冷的500 mL离心管中,冰上放置10 min;4℃,5 000 r·m-1离心15 min,用50 mL冰冷的EPWB缓冲液洗涤3次;用50 mL冰冷的EPB溶液洗涤1次;用2 mL冰冷的电转化缓冲液悬浮;分装每管100 μL。制备好的感受态细胞可以使用,并放入-70℃冰箱中待用。
将重组质粒pPG612-PLFN分别与电转化感受态细胞混匀后,冰上放置5 min;将其转入2 mm的预冷电转化杯(Bio-Rad公司)中;以Transformation Apparatus 165-2101电击,参数为电压2.5 kV,电击时间约为5 ms;电击后加入450 μL冰预冷的MRS培养基,混匀;将菌液转移至1.5 mL离心管中,冰上放置10 min;37℃厌氧培养2 h;取适量菌液涂布于含有 10 μg·mL-1Cm的MRS琼脂培养基上,37℃厌氧培养2~3 d。
1.7.3 阳性重组质粒的鉴定
分别于各平板上挑取单菌落,分别接种于含有10 μg·mL-1Cm的MRS 液体培养基中,37 ℃厌氧培养过夜后,按1.6方法提取质粒。对重组质粒进行Bam H I单酶切,Sac I、Xhol I双酶切鉴定,同时进行PCR鉴定。
2 结果与分析
2.1 重组质粒pMD18-T-PLFN的鉴定
如图1可知,重组质粒pMD18-T-PLFN用Bam H I单酶切后得到大小约为3 600 bp的基因片段,用Sac I、Xhol I双酶切后得到大小约为1 000和2 600 bp基因片段,以P1、P2为引物进行PCR扩增,得到约为1 000 bp基因片段,与预计大小相符合。表明PLFN基因片段插入到克隆载体pMD18-T中。
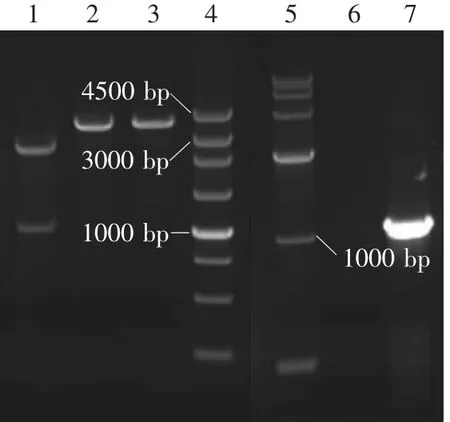
图1 pMD18-T-PLFN 重组质粒鉴定Fig.1 Identification of recombinant plasmid pMD18-T-PLFN
测序结果表明获得大小为1 077 bp的PLFN基因与预期相符,通过与GenBank上已发表的序列相比对,结果表明本研究获得的长白猪PLFN基因与GenBank 序列 AY306198.1、M81327.1、L77887.1及M92089.1的核苷酸同源性分别达99.16%,99.07%,99.63%和99.35%,可见PLFN基因在核苷酸序列上高度保守。
2.2 pPG612-PLFN基因干酪乳杆菌表达载体构建鉴定的结果
如图2所示,重组质粒pPG612-PLFN单酶切后得到大小为4 600 bp的片段,用SacⅠ、XholⅠ双酶切后得到大小约为1 000和3 600 bp片段。以P1、P2为引物进行PCR扩增,得到约为1 000 bp基因片段,与预计大小相符合。表明PLFN基因片段已插入到表达载体pPG612.1中。

图2 pPG611.1-Sa质粒的酶切及PCR鉴定Fig.2 Restriction enzyme and PCR analysis of recombinant plasmid pPG611.1-Sa
2.3 表达载体重组质粒的构建图谱
结果见图3。
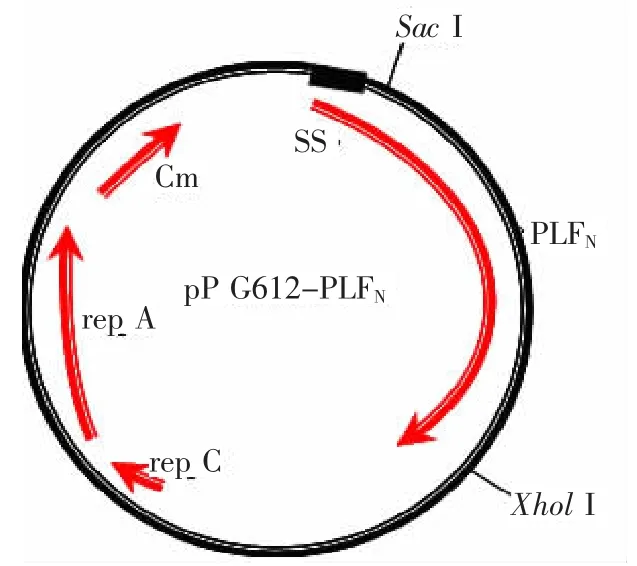
图3 重组质粒pPG612-PLFN 构建图谱Fig.3 Constructed map of recombinant plasmid pPG612-PLFN
序列测定结果表明,PLFN基因已成功插入到表达载体质粒pPG612.1的Sac I与Xhol I酶切位点之间,pPG612-PLFN/Lactobacillus casei 393为分泌表达的载体系统,表达蛋白将大部分分泌于培养菌体的上清液中。
2.4 重组乳酸菌的构建结果
将重组质粒pPG612-PLFN转化入各种干酪乳杆菌后,各重组乳酸菌的构建结果见图4~7。由图可见,PCR扩增均获得约1 000 bp的PLFN基因片段,Sac I、Xhol I双酶切分析均获得约1 000 bp的目的基因片段和约3 600 bp的表达载体片段,表明已获得转化了猪乳铁蛋白N端基因的四种重组乳酸菌,分别为Lactobacillus casei ATCC393,Lactobacillus plantarum KLDS1.0677,Lactobacillus paracasei KLDS1.0652,Lactobacillus pentosus KLDS 1.0413。
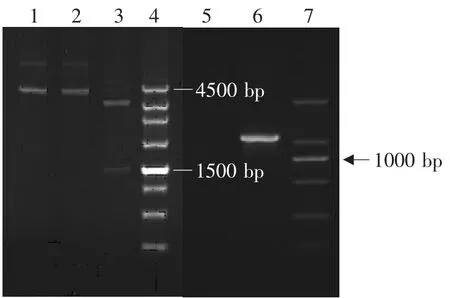
图4 重组副干酪乳杆菌KLDS1.0652鉴定Fig.4 Identification of recombinant Lactobacillus paracasei KLDS1.0652
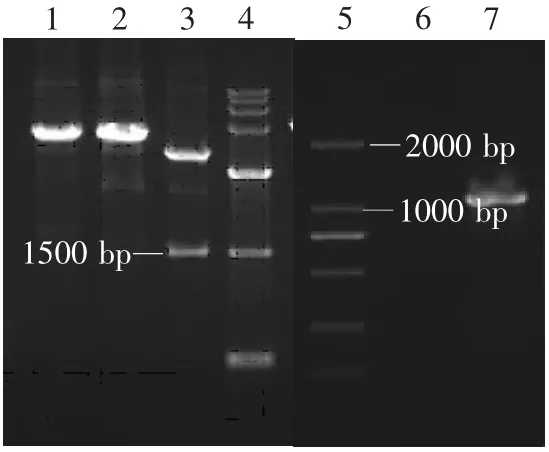
图5 重组植物乳杆菌KLDS1.0344鉴定Fig.5 Identification of recombinant Lactobacillus plantarum KLDS1.0344
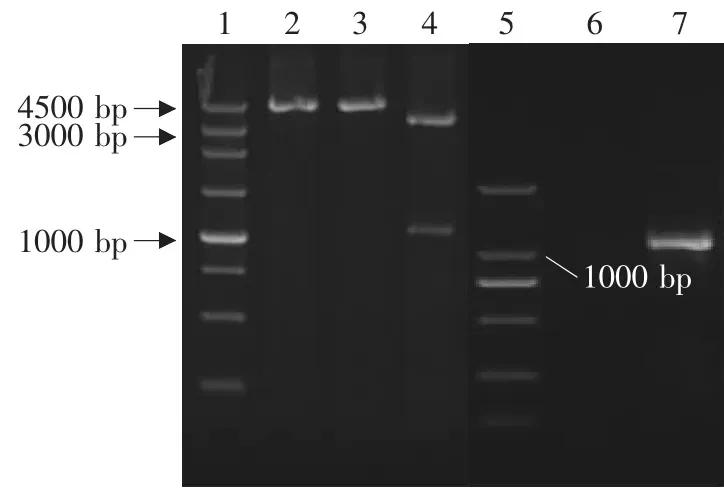
图6 重组干酪乳杆菌ATCC393鉴定Fig.6 Identification of recombinant Lactobacillus casei ATCC393
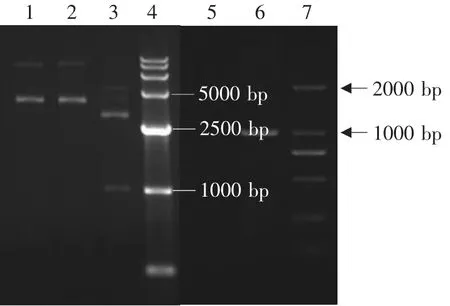
图7 重组戊糖乳杆菌KLDS1.0413鉴定Fig.7 Identification of recombinant Lactobacillus pentosus KLDS1.0413
3 讨论与结论
干酪乳杆菌作为表达和传递外源基因的载体系统具有益生性、安全性,以及可以通过先天性免疫激活肠道黏膜免疫系统,不会刺激体产生抗体等重要特点[9],尤其是分泌型载体表达的外源蛋白无需纯化,近年来受到越来越多的重视。在表达动物的病原基因作为黏膜免疫的口服疫苗以及一些安全级食品添加剂上日趋成熟。鉴于乳杆菌种类多(50多种),而不同乳杆菌生化特性、生态及分子构成的不同[10],本研究在乳酸菌的宿主菌选择上,除用了干酪乳杆菌ATCC393外,另选用了同是乳杆菌的植物乳杆菌、副干酪乳杆菌和戊糖乳杆菌。其中植物乳杆菌由于发酵断奶仔猪料具有适口性好、易消化吸收等优良特点,同时能给断奶仔猪提供一定的乳酸和有益菌,抑制有害菌的繁殖,从而可以缓解断奶腹泻和提高抗病力等特性[11-12]。Nemcova等报道副干酪乳杆菌可显著降低刚断奶幼猪体内梭菌属和肠杆菌的数量[13];已有研究表明[14],戊糖乳杆菌本身作为为活菌制剂,可抑制有害菌群的生长繁殖,促进有益菌在动物的肠道内定植,从而可防治仔猪腹泻。
乳铁蛋白是在多种哺乳动物如人类、猴、牛和猪的乳汁中含有的一种带正电荷的糖蛋白,是由一条带有两个铁离子结合区域的肽链折叠而成,分子量在不同动物有所区别,大约是72~85 ku之间。乳铁蛋白能够刺激机体产生系统性和黏膜免疫防御而抑制和杀死细菌,作为免疫刺激剂,乳铁蛋白可以改进免疫功能和加强宿主的防御系统,减少断乳仔猪疾病感染和断乳应激反应的发生[15-16]。Wu等的研究认为[17],乳铁蛋白可能是在家畜饲养上一种非常重要的抗生素替代品。猪乳铁蛋白研究者发现乳铁蛋白的许多功能都与乳铁蛋白N-叶密切相关,而且乳铁蛋白肽(Lctoferricin,Lfcin),被认为是乳铁蛋白的抗菌活性中心)也位于乳铁蛋白N-叶。Lfcin是乳铁蛋白在消化道内经酸性胃蛋白酶水解后的小肽[18],位于乳铁蛋白N-叶中的17~42氨基酸残基之间,具有广谱抗细菌、抗病毒和抗真菌活性,还是乳铁蛋白与淋巴细胞表面受休和细菌表面LPS结合的部位。因此为了在基因工程中有效表达猪乳铁蛋白,本研究选用猪乳铁蛋白N端基因序列作为靶基因,N端序列乳酸菌表达载体的成功构建,为研究猪乳铁蛋白的抗菌活性机制及近一步在体内应用发挥其广谱的抑菌、抗病毒、免疫调节作用等生物学功能奠定基础。
[1]汪以真,刘光富,初晓娜,等.猪乳铁蛋白N-叶基因PLF-N在大肠杆菌中的克隆、表达及特性分析[J].农业生物技术学报,2005,13(5):635-638.
[2]Chen H L,Lai Y W,Yen C C,et al.Production of recombinant porcine lactoferrin exhibiting antibacterial activity in methylotrophic yeast,Pichia pastoris[J].J Mol Microbiol Biotechnol,2004(8):141-149.
[3]Chen H L,Wang L C,Chang C H,et al.Recombinant porcine lactoferrin expressed in the milk of transgenic mice protects neonatal mice from a lethal challenge with enterovirus type 71[J].Vaccine,2008,26(7):891-898.
[4]Pecorini C,Savazzini F,Martino P A,et al.Heterologous expression of biologically active porcine lactoferrin in Pichia pastoris yeast[J].Vet Res Commun,2005,2:379-382.
[5]Wang S R,Lin T Y,Chen C M,et al.Isolation and expression of a porcine lactoferrin gene[J].Am J Vet Res,1997,58(10):1152-1158.
[6]Wang S H,Yang T S,Lin S M,et al.Expression,characterization,and purification of recombinant porcine lactoferrin in Pichia pastoris[J].Protein Expr Purif,2002,25(1):41-49.
[7]Anderson D G,Mckay L L.Simple and rapid method for isolating large plasmid DNA from lactic streptococci[J].Appl Environ Microbiol,1983,46:549-552.
[8]Holo H,Nes I F.High-frequency transformation,by electroporation,of Lactococcus lactis subsp.cremoris grown with glycine in osmotically.stabilized media[J].Appl Environ Microbiol,1989,55:3119-3123.
[9]Galdeano C M,Perdigón G.The probiotic bacterium Lactobacillus casei induces activation of the gut mucosal immune system through innate immunity[J].Clin Vaccine Immunol,2006,13(2):219-226.
[10]崔艳伟,孟祥晨.乳杆菌的分离与鉴定[J].东北农业大学学报,2008,39(6):115-119.
[11]李想,王然,程建军,等.植物乳杆菌培养基的优化[J].东北农业大学学报,2008,39(9):96-99.
[12]刘金萍,王士长.植物乳杆菌N3发酵断奶仔猪料对饲料pH值及菌群的影响[J].饲料工业,2009,30(17):16-18.
[13]Nemcova R,Bomba A,Gancarcikova S,et al.Study of effect of La-ctobacillus paracasei and Fructooligos accharides on the faecal micro-flora in weanling piglets[J]. Berliner Miilchner Tierarztl,1999, 112(6-7): 225-228.
[14]张丽芳,王占武.戊糖乳杆菌制剂防治仔猪腹泻效果初探[J].中国微生态学杂志,2008,20(12):568-569.
[15]Shan T,Wang Y,Wang Y,et al.Effect of dietary lactoferrin on the immune functions and serum iron level of weanling piglets[J].J Anim Sci,2007,85(9):2140-2146.
[16]Wang Y,Han F,Xu Z.Developmental gene expression of lactoferrin in duodenum and effect of weaning age on gene expression of lactoferrin in piglets[J].Arch Anim Nutr,2006,60(1):1-9.
[17]Wu S C,Chen H L,Yen C C,et al.Recombinant porcine lactoferrin expressed in the milk of transgenic mice enhances offspring growth performance[J].J Agric Food Chem.2007,55(12):4670-4677.
[18]Ikeda M,Nozaki A,Sugiyama K,et al.Characterization of antiviral activity of lactoferrin against hepatitis C virus infection in human cultured cells[J].Virus Res,2000,66:51-63.
