氧浓度变化对铜绿假单胞菌生物被膜生成的影响
2010-01-11崔冬清孙铁英黎建黄秀清
崔冬清孙铁英黎 建黄秀清
1中国医学科学院 北京协和医学院研究生院,北京 100730
2卫生部北京医院呼吸内科,北京 100730 3卫生部北京医院老年病研究所,北京 100730
氧浓度变化对铜绿假单胞菌生物被膜生成的影响
崔冬清1,孙铁英2,黎 建3,黄秀清3
1中国医学科学院 北京协和医学院研究生院,北京 100730
2卫生部北京医院呼吸内科,北京 1007303卫生部北京医院老年病研究所,北京 100730
目的研究氧浓度变化对铜绿假单胞菌生物被膜产生量的影响,探讨密度感应系统在生物被膜调控中的作用以及Ⅲ型分泌系统表达和生物被膜形成的关系。方法将 23株铜绿假单胞菌临床菌株在不同氧浓度环境下连续培养3 d,检测生物被膜和藻酸盐产生量,银染色法观察 ATCC27853产生胞外多糖的时间窗,实时定量多聚酶链反应(PCR)测定密度感应系统中调控基因LasI和RhlI的表达,Western blot方法观察Ⅲ型分泌系统的代表毒素胞外酶 S分泌情况。结果铜绿假单胞菌的生物被膜产生量 (R=0.455,P=0.000)及藻酸盐产生量 (R=0.367,P=0.000)均与氧浓度变化呈显著正相关。银染色法结果显示,高氧条件下胞外多糖合成时间缩短。实时定量 PCR结果显示,LasI、RhlI的相对表达量与氧浓度 (R=0.025,P=0.794;R=-0.044,P=0.653)、生物被膜产生量 (R=0.001,P=0.990;R=0.011,P=0.909)和藻酸盐产生量 (R=0.029,P=0.770;R=0.193,P=0.064)间无明显相关性。Western blot结果表明,铜绿假单胞菌在氧浓度为 10%~30%条件下可分泌胞外酶 S,在高氧条件下不分泌胞外酶 S。结论高氧可促进铜绿假单胞菌生物被膜的产生。Las系统和 Rhl系统可能因为菌群数量处于低水平而未在生物被膜形成早期阶段参与生物被膜的调控过程。生物被膜的产生增多可能抑制Ⅲ型分泌系统的表达,使该菌毒力发挥受到限制。
铜绿假单胞菌;生物被膜;氧浓度;密度感应系统;Ⅲ型分泌系统
DO I:10.3881/j.issn.1000-503X.2010.03.015
铜绿假单胞菌是院内获得性感染的重要致病菌之一,流行病学统计资料表明该菌院内获得性感染发病率远高于社区获得性感染[1-3],推测其原因可能与院内患者免疫力低下及使用广谱抗生素有关[4]。然而,单纯的外源性因素并不能完全解释铜绿假单胞菌院内感染高发的原因,可产生生物被膜被认为是铜绿假单胞菌耐药和逃避人体免疫攻击的重要手段。藻酸盐是生物被膜的重要组成成分,具有抑制中性粒细胞活性、保护铜绿假单胞菌的作用[5]。密度感应系统是细菌协调群体生物学效应的一种有效手段,但其在生物被膜形成中的调控作用目前争议颇多。Ⅲ型分泌系统是铜绿假单胞菌主要的毒力因素之一,在急性感染中可发挥重要作用,对于Ⅲ型分泌系统和生物被膜形成这两种生物学行为的关系目前学术界也存在一些分歧。本研究观察了氧浓度变化对铜绿假单胞菌生物被膜及藻酸盐产生量的影响,探讨了密度感应系统在生物被膜调控中的作用以及Ⅲ型分泌系统表达和生物被膜形成的关系。
材料和方法
铜绿假单胞菌菌株的来源 23株铜绿假单胞菌临床株和标准菌株 ATCC27853由卫生部北京医院检验科细菌室提供,临床株均为呼吸内科等临床科室住院患者痰标本培养分离得来。所有菌株均经随机扩增 DNA多态性基因分型法 (random amplified polymorphic DNA,RAPD)证实来源于不同克隆[6]。
菌株培养 采用改良的微板法培养菌株,具体为:过夜培养的菌液经分光光度计 (UV-1601,日本岛津)测定 570 nm吸光度,LB培养液调定菌液浓度为 0.1OD,24孔培养板中每孔加入 200μl菌液、1张无菌特氟龙滤膜 (PTFE,北京北化黎明分离技术公司)和 1 ml LB培养液,放入密闭培养箱中连续培养 3 d[7]。每次培养持续通入 1种氧浓度的混合气体 (含有氧气和氮气),氧气浓度分别为 0、10%、20%、30%、40%、50%、60%。
被膜菌总蛋白量的测定 细菌连续培养 3 d后,PBS清洗滤膜上的浮游菌,加入细胞裂解液裂解细菌 20 min,超声 6 s,12 000 g、4℃低温离心 15 min,取上清用 Bradford法检测细菌总蛋白量。
生物被膜产生量的检测 细菌连续培养 3 d后,PBS清洗滤膜上的浮游菌,结晶紫染液 (法国梅里埃公司)染色滤膜 20 min,去离子水漂洗后加入 95%酒精脱色 10 min,脱色液用分光光度计测定 570 nm的吸光度,再用每株菌的总蛋白量校正生物被膜吸光度。
藻酸盐产生量的测定 细菌连续培养 3 d后,PBS清洗滤膜上的浮游菌,加入 1.2 ml硫酸 /硼酸钠混合物,100℃煮沸 5 min,加入 20μl 1%对-羟基联苯溶液显色,分光光度计测定 520 nm吸光度,通过藻酸盐 (美国 Sigma公司)标准曲线换算黏附在滤膜上的藻酸盐量,即为每株菌产生的藻酸盐总量,最后用被膜菌总蛋白量校正。
不同氧浓度下菌株 ATCC27853胞外多糖产生时间窗的观察 过夜培养菌株 ATCC27853,LB培养液调定菌液浓度至 0.1OD值。6孔培养板中每孔加入3 ml LB培养液和 1张无菌盖玻片 (20 mm×20 mm×0.7 mm),并加入 200μl菌液放入 37℃孵箱孵育,每次持续通入含 1种氧浓度的混合气体。每隔 1 h取出 1张盖玻片进行 5%硝酸银溶液染色。光学显微镜下若见灰黑色斑片状,则可鉴定为细菌合成的胞外多糖。
不同氧浓度下 LasI和 RhlI表达的测定 将 23株铜绿假单胞菌临床株以相同浓度的初始菌液分别在不同氧浓度环境下连续孵育 8 h,离心,留沉淀。采用 Trizol(美国 Invitrogen公司)法提取 RNA,再反转录为 cDNA,以管家基因Rpsl为内对照,荧光实时定量聚合酶链反应 (Real time polymerase chain reaction,Real time PCR)检测LasI和RhlI表达。PCR体系、扩增参数参照 Power SYBR Green试剂盒 (美国 AB I公司)说明书。扩增LasI基因上游引物序列为 5-CAAGTTGCGTGCTCAAGTGT-3,下游序列为 5-GGGAAGGTGTTCTTCAGCAT-3;RhlI上游引物序列为 5-CTTGGTCATGATCGAATTGCTC-3,下游序列为5-ACGGCTGACGACCTCACAC-3;管家基因Rpsl上游引物序列为 5-CGGCACTGCGTAAGGTATGC-3,下游序列为 5-CCCGGAAGGTCCTTTACAC-3。采用相对定量公式 2-ΔΔCt计算各目的基因LasI和RhlI的 mRNA相对表达量:目的基因的量 =2-ΔΔCt,其中 Δ ΔCt=ΔCt(目的基因)-ΔCt(管家基因Rpsl)。
不同氧浓度下胞外酶 S分泌的测定 将 23株铜绿假单胞菌临床株以相同浓度的初始菌液分别在不同氧浓度环境下连续孵育8 h,离心,取上清。蛋白经高温变性后,采用聚丙烯酰胺凝胶电泳分离约 1 h,300 V、低温 1 h转至 PVDF膜,室温封闭 1 h。分别与 1∶2 500鸡抗 Exoenzyme S单克隆抗体 (英国 Abcam公司)4℃孵育过夜,漂洗后加入 1∶2 500 HRP标记的兔抗鸡 IgG相关抗原抗体 (北京友谊中联公司),室温孵育 1 h,漂洗后于暗室中加入混合 ECL发光缓冲液,压 X-Ray胶片,显影、定影、胶片扫描。
统计学处理 采用 SPSS13.0统计软件,相关性分析采用 Spearman检验,组间差异性分析采用双向方差分析,P<0.05为差异有统计学意义。
结 果
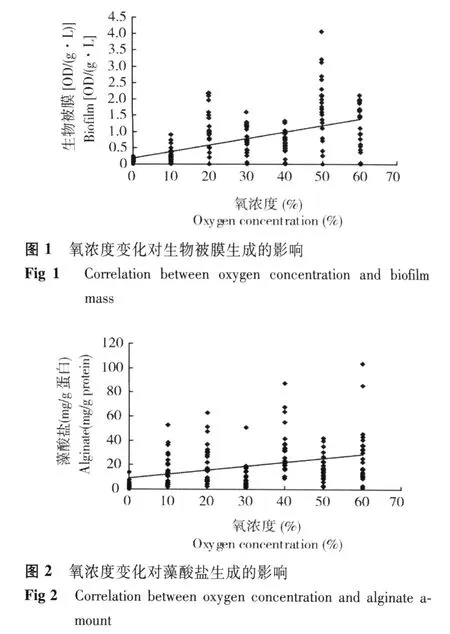
不同氧浓度条件下铜绿假单胞菌生物被膜产生量 当氧浓度为 50%时,生物被膜产生量达最高值[4.05 OD/(g·L)];趋势性分析结果表明,随着氧浓度升高,生物被膜产生量逐渐上升 (图 1)。相关性分析结果显示,铜绿假单胞菌的生物被膜产生量与氧浓度呈显著正相关 (R=0.455,P=0.000)。差异性分析结果显示,不同氧浓度下铜绿假单胞菌的生物被膜产生量间差异有统计学意义 (F=3.934,P=0.000)。
不同氧浓度条件下铜绿假单胞菌藻酸盐产生量当氧浓度为 60%时,藻酸盐产生量达最高值(85.22 mg/g蛋白);趋势性分析结果表明,随着氧浓度升高,藻酸盐产生量逐渐升高 (图 2)。相关性分析结果显示,铜绿假单胞菌的藻酸盐产生量与氧浓度呈显著正相关 (R=0.367,P=0.000)。差异性分析结果显示,不同氧浓度下铜绿假单胞菌的藻酸盐产生量间差异有统计学意义 (F=5.905,P=0.000)。
不同氧浓度条件下 ATCC27853产生胞外多糖的时间窗 氧浓度为 40%、50%或 60%时,ATCC27853培养 1 h即可出现银染的胞外多糖;氧浓度分别为20%、30%时,菌株需培养 2 h以上才出现胞外多糖;氧浓度为 10%时,胞外多糖最早出现时间为 4 h;无氧培养条件下,菌株培养 8 h出现胞外多糖。
LasI和 RhlI表达与氧浓度、生物被膜产生量和藻酸盐产生量之间的关系 相关性分析结果表明,LasI的相对表达量与氧浓度 (R=0.025,P=0.794)、生物被膜产生量 (R=0.001,P=0.990)和藻酸盐产生量 (R=0.029,P=0.770)间无显著相关性;RhlI的相对表达量与氧浓度 (R=-0.044,P=0.653)、生物被膜产生量 (R=0.011,P=0.909)和藻酸盐产生量 (R=0.193,P=0.064)间也无显著相关性。
不同氧浓度下胞外酶 S的分泌情况 当氧浓度为 10%时,菌株 15、16能够分泌胞外酶 S(图3A);在氧浓度为 20%时,菌株 1~5、7~10、15、16、19、23能够分泌胞外酶 S(图 3B、C、D);当氧浓度为 30%时,菌株 1、2、4、5、9、10、15能够分泌胞外酶 S(图 3E、F);当氧浓度为 0、40%、50%和 60%时,无菌株分泌胞外酶 S。
讨 论
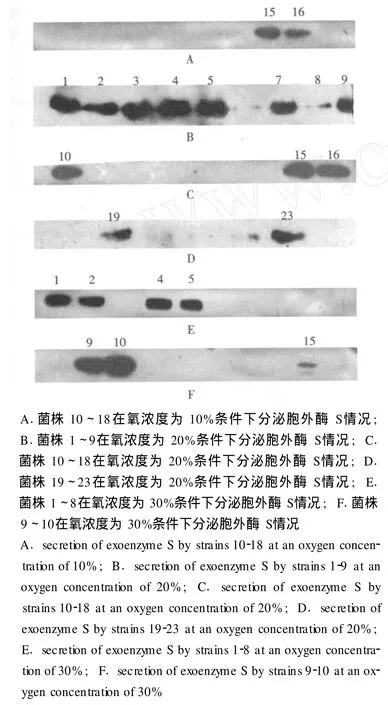
图 3 不同氧浓度下胞外酶 S的分泌情况Fig 3 The secretions of exoenzyme S under different levels of environmental oxygen
生物被膜是细菌分泌的一种多糖复合物,覆盖在细菌表面,可保护细菌免受人体免疫细胞的攻击,并可通过限制抗生素的渗透等作用提高细菌耐药性。细菌在人体内产生生物被膜,导致细菌不易被清除而造成的反复感染被称为生物被膜病,目前临床上对于生物被膜病的治疗方法十分有限[5]。藻酸盐是生物被膜的重要组成成分,可增强细菌黏附性,促进生物被膜形成,提高细菌的耐药性;并可直接抑制中性粒细胞和巨噬细胞的趋化及吞噬作用,对细菌同样能起到保护作用[8]。
铜绿假单胞菌以往被认为是需氧菌。1986年,Bazylinski等[9]发现该菌在无氧条件下可利用含氮化合物进行脱氮作用产能并生长。Worlitzsch等[10]对比了在无氧和空气氧条件下铜绿假单胞菌产生藻酸盐量的差异,发现铜绿假单胞菌在无氧条件下产生的藻酸盐更多。本研究则观察了不同浓度氧对铜绿假单胞菌生物被膜及藻酸盐产生量的影响,结果显示,铜绿假单胞菌的生物被膜产生量及藻酸盐产生量均与氧浓度变化呈显著正相关;当氧浓度为 50%时,生物被膜产生量达最高值;当氧浓度为 60%时,藻酸盐产生量达最高值;提示高氧环境促进了生物被膜和藻酸盐的产生。本研究中使用的高氧浓度分别为 30%、40%、50%、60%,最高氧浓度之所以选择 60%,是因为临床上氧疗的安全范围低于 60%,更高浓度的氧疗容易导致氧中毒。此外,本研究还采用银染色法光镜下观察了不同氧浓度条件下铜绿假单胞菌生成胞外多糖的时间窗,结果发现,越高氧浓度环境下合成胞外多糖所需要的时间越短,提示高氧可促进胞外多糖的合成,有利于生物被膜的形成。
为了探查密度感应系统在生物被膜形成中的作用,本研究采用实时定量 PCR方法测定不同氧浓度条件下铜绿假单胞菌临床株 Las系统和 Rhl系统中信号分子合成酶基因LasI和RhlI的表达情况,结果显示LasI和RhlI的相对表达量与对应菌株在同一条件下生物被膜和藻酸盐产生量无显著相关性。分析其原因可能是本研究是将 23株临床菌株培养 8 h后提取 RNA、反转录,再进行实时定量 PCR测定目的基因表达,由于 8 h的时间段是处于生物被膜形成过程的早期,而密度感应系统作用的发挥需要有一定数量菌群才能产生级联放大效应,所以对于生物被膜形成的早期,由于菌体数量处于低水平,此时密度感应系统可能不发挥主要作用。
为了观察Ⅲ型分泌系统和生物被膜产生量的关系,本研究在不同氧浓度环境下培养了铜绿假单胞菌临床株,并分离上清,检测菌株分泌Ⅲ型分泌系统的代表毒素——胞外酶 S的能力,结果显示,23株铜绿假单胞菌临床株只有在氧浓度为 10%~30%的条件下分泌胞外酶 S。推测低氧环境下可能菌体处于低代谢状态,导致胞外酶 S分泌下调;而高氧环境虽然有利于生物被膜产生,但并不利于胞外酶 S分泌,提示当铜绿假单胞菌大量合成生物被膜时,Ⅲ型分泌系统表达受到抑制,侵袭力下降。
综上所述,本研究结果显示高氧环境有利于铜绿假单胞菌生长及其生物被膜和藻酸盐产生,故笔者推测医源性氧疗可能促进了铜绿假单胞菌产生生物被膜和藻酸盐,导致该菌在人体内的耐药和寄植,而这可能就是铜绿假单胞菌院内高感染的原因之一。建议针对有铜绿假单胞菌感染高危因素的患者应避免过高浓度的氧疗,以免造成有利于铜绿假单胞菌寄植的环境。
[1] WeberDJ,Rutala WA,Sickbert-Bennett EE,et al.Microbiology of ventilator-associated pneumonia compared with that of hospital-acquired pneumonia[J].Infect Control Hosp Epidemiol,2007,28(7):825-831.
[2] DriscollJA,Brody SL,KollefMH.The epidemiology,pathogenesis and treatment of pseudomonas aeruginosa infections[J].Drugs,2007,67(3):351-368.
[3] Kollef MH,Shorr A,Tabak YP,et al.Epidemiology and outcomes of health-care-associated pneumonia:results from a large US database of culture-positive pneumonia[J].Chest,2005,128(6):3854-3862
[4] Koulenti D,Rello J.Gram-negative bacterial pneumonia:aetiology and management[J].Curr Opin Pulm Med,2006,12(3):198-204.
[5] Kobayashi H.Airway biofilms:implications for pathogenesis and therapy of respiratory tract infections[J].Treat Respir Med,2005,4(4):241-253.
[6] Giske CG,Libisch B,Colinon C,et al.Establishing clonal relationships between V IM-1-like metallo-beta-lactamase-producing Pseudomonas aeruginosa strains from four European countries by multilocus sequence typing[J].J Clin Microbiol,2006,44(12):4309-4315.
[7] Knezevic P,Petrovic O.A colorimetric microtiter plate method for assessment of phage effect on Pseudomonas aeruginosa biofilm[J].J MicrobiolMethods,2008,74(2-3):114-118.
[8] Leid JG,W illson CJ,Shirtliff ME,et al.The exopolysaccharide alginate protects Pseudomonas aeruginosa biofilm bacteria from IFN-gamma-mediated macrophage killing[J].J Immunol,2005,175(11):7512-7518.
[9] BazylinskiDA,Soohoo CK,Hollocher TC.Growth of Pseudomonas aeruginosa on nitrous oxide[J].Appl EnvironMicrobiol,1986,51(6):1239-1246.
[10] Worlitzsch D,Tarran R,UlrichM,et al.Effects of reduced mucus oxygen concentration in airway Pseudomonas infections of cystic fibrosis patients[J].J Clin Invest,2002,109(3):317-325.
Effect of D ifferentLevels of Environmental Oxygen on the Biofilm Production ofPseudom onas aeruginosa
CU IDong-qing1,SUN Tie-ying2,L I Jian3,HUANG Xiu-qing3
1Graduate School,CAMS and PUMC,Beijing 100730,China2Department of RespiratoryMedicine,Beijing Hospital,Ministry of Health,Beijing 100730,China3National Institute of Geriatrics,Beijing Hospital,Ministry of Health,Beijing 100730,China
SUN Tie-ying Tel:010-85136242,E-mail:suntieying3@hotmail.com
ObjectiveTo investigate the relationship among oxygen concentration,quorum sensing system,typeⅢsecretion system,and biofilm production ofPseudomonas aeruginosa.M ethods A total of 23 clinical strains ofPseudomonas aeruginosawere cultured at different levels of environmental oxygen for three days.Then biofilm mass and alginate were quantified.The expression levels ofLasIandRhlIwere detected by real time polymerase chain reaction(PCR).The secretion of exoenzyme Swas examined byWestern blot.ResultsBoth the biofilm mass(R=0.455,P=0.000)and alginate(R=0.367,P=0.000)were positively correlated with oxygen concentration.Real time PCR showed that the expression levels ofLasIandRhlIwere not significantly correlated with oxygen concentration(R=0.025,P=0.794;R=-0.044,P=0.653),the production of biofilm(R=0.001,P=0.990;R=0.011,P=0.909),or alginate(R=0.029,P=0.770;R=0.193,P=0.064).Western blot showed that the optimal oxygen concentration range for exoenzyme S secretion ofPseudom onas aeruginosaranged 10%to 30%.Conclusions Hyperoxia can promote the production of biofilm and alginate byPseudom onas aeruginosa.Las/Rhl system may not participate in biofilm production at the early stage due to the low bacteria amount.The increased production of biofilm may inhibit the expression of TypeⅢSecretion system and thus inhibit bacterial virulence.
Pseudomonas aeruginosa;biofilm;oxygen concentration;quorum sensing system;typeⅢsecretion system
Acta Acad M ed Sin,2010,32(3):310-314
孙铁英 电话:010-85136242,电子邮件:suntieying3@hotmail.com
R378.99+1
A
1000-503X(2010)03-0310-05
2009-05-19)
·论 著·
