人脂联素基因的克隆及序列比较分析
2009-11-29陈团生郑金贵
刘 峰 陈团生 郑金贵
(福建农林大学生命科学学院,福建 福州 350002)(福建农林大学生命科学学院,福建 福州 350002; 福建农林大学医院,福建 福州 350002)(福建农林大学生命科学学院,福建 福州 350002)
人脂联素基因的克隆及序列比较分析
刘 峰 陈团生 郑金贵
(福建农林大学生命科学学院,福建 福州 350002)(福建农林大学生命科学学院,福建 福州 350002; 福建农林大学医院,福建 福州 350002)(福建农林大学生命科学学院,福建 福州 350002)
目的:克隆人脂联素基因,为进一步重组蛋白表达和生物活性等基础研究与临床应用奠定基础。方法:应用RT-PCR法自人大网膜脂肪组织的总RNA中克隆人脂联素全长基因,采用T-A克隆法重组到pMD18-T中,通过菌液PCR鉴定阳性克隆后,测序鉴定。并采用聚类分析等对其序列进行比较分析。结果:克隆人脂联素基因全长为735bp,并登录GenBank(登录号:EU420013);其编码244个氨基酸,理论分子量为26 413.64Da,预测的等电点为5.42。邻接法聚类分析表明,人脂联素与已报道的其它哺乳动物的脂联素相似性达83%~96%。结论:成功地克隆了人脂联素基因全长序列,人脂联素与已报道的其它哺乳动物的脂联素序列具有很高的同源性。
人脂联素;基因克隆;序列分析
近年来的研究表明,糖尿病患者血液中脂联素(Adiponectin)水平降低会导致其对胰岛素敏感性的下降[1]。脂联素是Scherer等[2]发现的一种由脂肪组织特异性分泌的约30kDa的脂肪细胞因子[3],能有效促进脂肪酸氧化和减少糖异生,从而降低血糖和血脂水平[4,5]。对于2型糖尿病及心血管疾病患者,其血浆中的脂联素水平远远低于正常人,且通常伴有胰岛素抵抗和高胰岛素血症。而增加脂联素水平则能显著增强胰岛素的敏感性,并能抑制肝糖元生成[6-8]。此外,脂联素水平的下降也是导致动脉粥样硬化的重要因素[9];利用转基因技术在机体高表达脂联素后可明显对抗ApoE基因缺陷小鼠动脉粥样硬化的形成[8]。脂联素水平的降低还可致血管功能下降以及冠心病发病危险性增加[1,9,10]。本研究从人大网膜脂肪组织克隆脂联素编码区全长基因,并对其进行了序列比较分析,为进一步的重组蛋白表达和生物活性等理论研究与临床应用奠定基础。
1 材料与方法
1.1材料取自于某省立医院胃肠外科胃溃疡患者行胃部分切除术中取出的大网膜脂肪组织,迅速将其装入mRNA保护液(大连宝生物公司),-80℃低温冰箱保存备用。
1.2菌种、质粒与主要试剂大肠杆菌(E. coli)DH5α由实验室保存提供;克隆载体pMD18-T、ExTaq酶、PrimeScriptTM1st Strand cDNA Synthesis Kit、各种限制性内切酶、T4DNA连接酶等购自大连宝生物(TaKaRa)公司。
1.3人大网膜脂肪组织总RNA的提取采用Qiagen公司RNAeasy mini kit提取人大网膜脂肪组织的总RNA;各种RNase-free的枪头、离心管、RNase-free超纯水均购置于百泰克生物工程公司。
1.4RT-PCR使用PrimeScriptTM 1st Strand cDNA Synthesis Kit(TaKaRa)将1μg的总RNA反转录成cDNA。以Maeda等[11]报道的人脂联素基因序列为基础设计出一对特异性引物(划线部位为增加的BamHI与SacI酶切位点):
以反转录获得的cDNA为模板进行PCR反应。扩增条件为94℃ 5min,35个循环(94℃ 1min,56℃ 5min,72℃ 1min),72℃ 10min。
1.5克隆载体构建RT-PCR产物胶回收操作步骤参照百泰克生物公司多功能DNA纯化回收试剂盒的说明进行。参照TaKaRa公司pMD18-T Vector试剂盒的说明将目的片段连接到pMD18-T Vector上。转化大肠杆菌DH5α,筛选抗Amp菌落,同时进行菌落PCR鉴定和测序,并命名为pMD-ADPN。
1.6克隆产物酶切鉴定重组质粒pMD-ADPN的提取参照百泰克生物公司质粒小量提取纯化试剂盒的说明进行。采用Bam HI、Sac I双酶切pMD-ADPN,对其进行鉴定。
1.7脂联素氨基酸序列比对、进化树的构建使用CLUSTALX 2.0程序将人脂联素的氨基酸序列与其它哺乳动物脂联素的氨基酸进行多序列比对(其它哺乳动物的脂联素氨基酸序列分别来源于GenBank、EMBL和PDB数据库等)。使用Prosite软件分析其功能基团;利用MEGA 4.0程序,采用邻接法(1 000 replicates; seed = 64 238)聚类分析构建系统进化树。
2 结 果
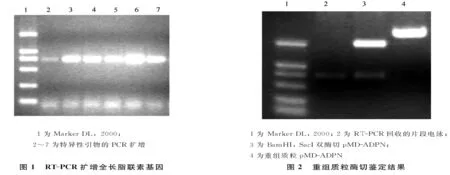
2.1RT-PCR结果应用Qiagen公司RNAeasy mini kit提取人大网膜脂肪组织获得高质量的总RNA;采用PrimeScriptTM1st Strand cDNA Synthesis Kit将总RNA转录成cDNA后,RT-PCR结果显示在750bp左右出现目标条带(见图1)。
2.2克隆产物酶切鉴定图谱采用T-A克隆方法将PCR产物与pMD18-T连接、转化DH5α;提取质粒pMD-ADPN;酶切鉴定表明重组质粒为目的质粒(见图2)。
2.3测序结果测序表明,该cDNA全长为735bp,为一个完整的开放阅读框ORF(见图3);ORF编码244个氨基酸残基,未修饰蛋白的理论分子量为26 413.64Da,预测的等电点为5.42。我们克隆的人脂联素基因的全长cDNA已在GenBank 登录,登录号为EU420013。

图3人脂联素基因全长cDNA及其推导的氨基酸序列
2.4氨基酸序列比较与分析将我们克隆的脂联素与已报道的其它哺乳动物的脂联素进行同源性分析,结果表明其与兔(Oryctolaguscuniculus,Oc-Acrp30:ABC60052)、狗(Canislupusfamiliaris,Clf-Acrp30: XP_535838)、灰狼(Canislupus,Cl-Acrp30: ACI01768)、家猫(Feliscatus,Fc-Acrp30: NP_001078907)、狸(Nyctereutesprocyonoides,Np-Acrp30: AAX73246)、欧洲棕熊(Ursusarctos,Ua-Acrp30: ACI01763)、北极狐(Vulpeslagopus,Vl-Acrp30: AAX73247)、欧洲臭鼬(Mustelaputorius,Mp-Acrp30: ACI01766)、挪威鼠(Rattusnorvegicus,Rn-Acrp30: NP_653345)、紫貂(Marteszibellina,Mz-Acrp30: ACI01764)、小家鼠(Musmusculus,Mm-Acrp30: AAH28770)、牛(Bostaurus, Bt-Acrp30: AAK58902)、猕猴(Macacamulatta,Mmu-Acrp30: AF404407)、马(Equuscaballus, Ec-Acrp30: XP_001499564)、水貂(Neovisonvison,Nv-Acrp30: ACI01765)、猿猴(Macacafuscata,Mf-Acrp30: BAG16752)等脂联素的序列同源性都很高,达83%~96%的一致性。结果如图4所示(见第77页彩色图版)。

一致的序列用点标出;横线表示空缺。人脂联素的氨基酸序列下标红线。哺乳动物脂联素氨基酸序列与人脂联素的氨基酸序列的相似性被列在序列比对的末端。GenBank等数据库登录号分别为:兔(Oryctolagus cuniculus, ABC60052)、狗(Canis lupus familiaris, XP_535838、灰狼 (Canis lupus,ACI01768)、家猫(Felis catus, NP_001078907)、狸(Nyctereutes procyonoides, AAX73246)、欧洲棕熊(Ursus arctos, ACI01763)、北极狐(Vulpes lagopus, AAX73247)、欧洲臭鼬(Mustela putorius,ACI01766)、紫貂(Martes zibellina, ACI01764)、小家鼠(Mus musculus, AAH28770)、牛(Bos taurus, AAK58902) 、猕猴(Macaca mulatta, AF404407) 、马(Equus caballus,XP_001499564)、水貂(Neovison vison, ACI01765)、猿猴(Macaca fuscata,BAG16752)。图4 使用GLUSTAL X 2.0将人脂联素的氨基酸序列与其他哺乳动物的脂联素氨基酸进行比对
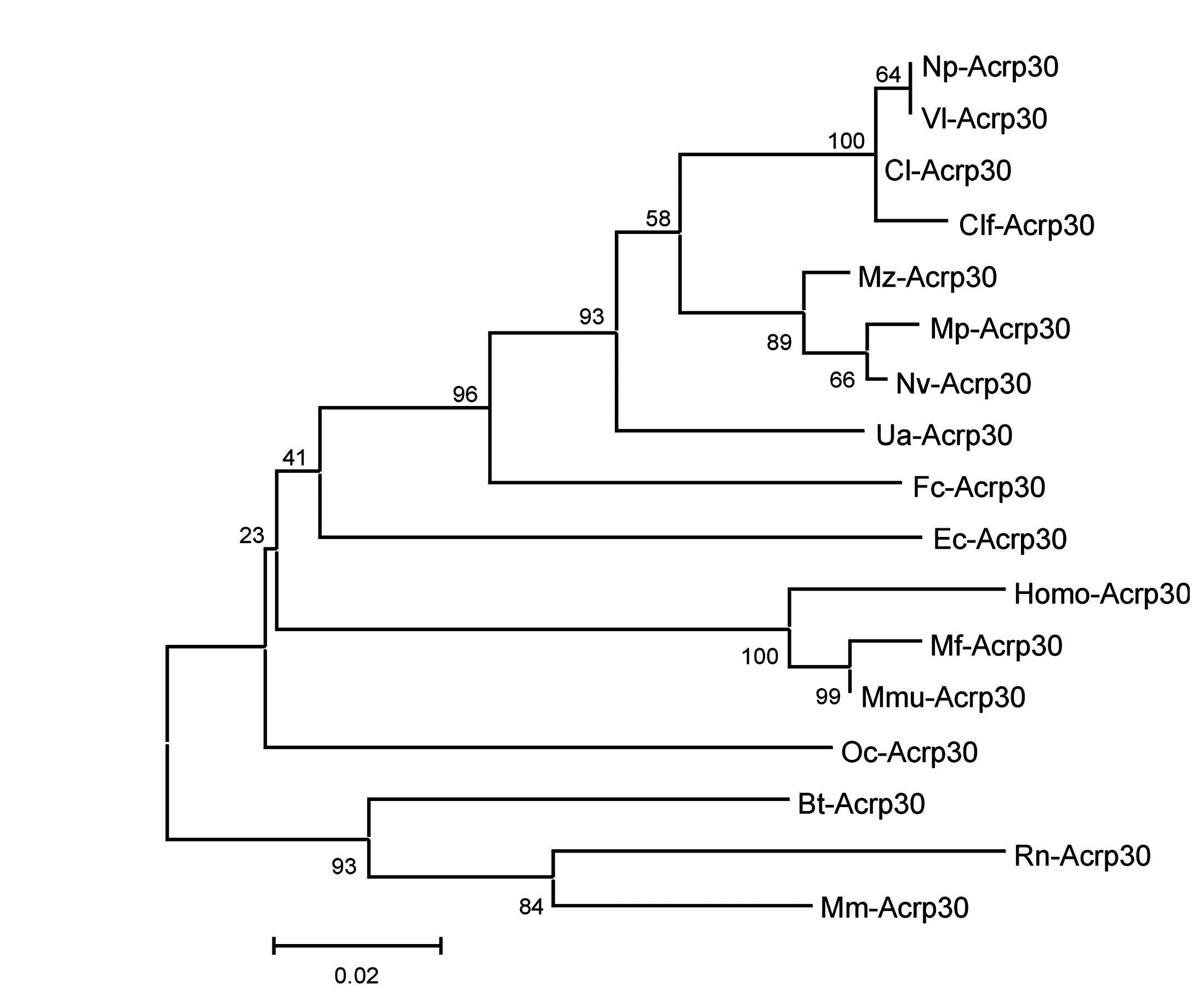
图5 利用MEGA 4.0程序,采用邻接法(1 000 replicates;seed=64 238)聚类分析构建哺乳动物17个种的系统进化树
利用蛋白质功能位点分析软件Prosite对人脂联素氨基酸序列进行分析,人体的脂联素由244个氨基酸组成,包括氨基端信号肽序列(1-18aa)、胶原样结构域(19-107aa)和羧基末端的球形结构域等;高度保守的motifs包括GEKGEKGD、GPKGD、GRKGE等,对脂联素发挥活性功能起重要作用,组成了该家族的信号模式;其中胶原结构域内的四个高度保守的赖氨酸是蛋白糖基化及羟基化的位点。采用邻接法(neighbour joining,NJ)聚类分析结果表明,人脂联素与大多数哺乳动物的亲缘关系都较近(见图5)。
3 讨 论
人脂联素基因位于染色体3q27,该位点是2型糖尿病、代谢综合症和冠状动脉粥样硬化性心脏病(冠心病)的易感位点。2型糖尿病多在35~40岁之后发病,目前占我国糖尿病患者90%以上。2型糖尿病病人体内并非完全丧失产生胰岛素的能力,甚至有的患者体内胰岛素产生过多,因此应用胰岛素进行治疗的效果被大打折扣,即我们常说的“胰岛素抵抗”。研究表明胰岛素敏感性的下降是由于血中脂联素水平降低所致[1];当敲除脂联素基因后,高脂饮食可以诱发出机体严重的胰岛素抵抗[11]。对于脂联素基因敲除的小鼠,用压力负荷方式极易诱导出心肌肥厚,而将腺病毒带的脂联素基因重新转入体内后可以逆转这种趋势[9]。
脂联素对机体胰岛素的增敏可能是通过靶器官肝和肌肉而起作用的[12]。脂联素可通过加强胰岛素介导的胰岛素受体酪氨酸磷酸化,从而激活胰岛素受体底物-1(IRS-1)介导的胰岛素信号系统,使肌肉组织中糖摄取增加;通过激活5’AMP激活的蛋白激酶(AMPK)直接刺激葡萄糖利用和脂肪酸氧化,调节胰岛素敏感性[6, 13]。此外,脂联素抑制糖异生途径的两个关键酶基因(葡萄糖-6-磷酸酶及磷酸烯醇式丙酮酸羧激酶基因)的表达,从而降低血糖浓度,进而改善胰岛素抵抗[4, 6]。而脂联素水平低下可降低肌肉脂肪酸转运蛋白-1的mRNA和胰岛素受体底物-1(IRS-1)介导的胰岛素信号系统,从而导致严重的胰岛素抵抗[12]。提升脂联素水平能增强胰岛素的敏感性,并能降低血糖水平,因而在临床治疗上具有潜在的较大的价值。
本研究从人大网膜脂肪组织中提取总RNA,进行RT-PCR反应,获得含人脂联素基因的重组质粒载体pMD-ADPN。测序表明,与Maeda等[11]报道的人脂联素基因序列同源性达100%;未发生任何突变等,提示我们成功的克隆获得了目的基因。RNA的获取是本实验研究中一个基本关键环节。为了能够获得完整的、丰度较高的mRNA,我们在手术台获取人大网膜脂肪组织后,迅速将其装入mRNA保护液(TaKaRa),以防止RNA的降解。在提取过程中则严格防止RNase的污染。实验表明,通过使用Qiagen公司RNAeasy mini kit获得了高丰度和高质量的RNA,为本研究的顺利进行奠定了重要的基础。通过成功的克隆人脂联素基因,为重组表达脂联素,进一步为基础研究与临床应用提供了实验基础。
[1]Hotta K, Funahashi T, Bodkin N L,etal.Circulating concentrations of the adipocyte protein adiponectin are decreased in parallel with reduced insulin sensitivity during the progression to type 2 diabetes in rhesus monkeys[J]. Diabetes,2001,50 (5):1126-1133.
[2]Scherer PE,Williams S,Fogliano M,etal.A novel serum protein similar to C1q,produced exclusively in adipocytes[J]. Biol Chem,1995,270 (45):26746-26749.
[3]Fu Y,Luo N,Klein RL,etal.Adiponectin promotes adipocyte differentiation,insulin sensitivity,and lipid accumulation[J]. J Lipid Res,2005,46:1369-1379.
[4]Zhou H,Song X,Briggs M,etal.Adiponectin represses gluconeogenesis independent of insulin in hepatocytes[J]. Biochem Biophys Res Commun,2005,338:793-799.
[5]Heilbronm LK,Smith SR,Ravussin E. The insulin sensitizing role of the fat derived hormone adiponectin[J]. Curr Pharm Des,2003,9:1411-1418.
[6]Berg AH,Combs TP,Du X,etal.The adipocyte-secreted protein Acrp30 enhances hepatic insulin action[J]. Nat Med,2001,7:947-953.
[7]Bacha F,Saad R,Gungor N,etal.Adiponectin in youth:relationship to visceral adiposity,insulin sensitivity,and beta-cell function[J]. Diabetes Care,2004,27(2):547-552.
[8]Yamauchi T,Kamon J,Ito Y,etal.Cloning of adiponectin receptors that mediate anti diabetic metabolic efects[J]. Nature,2003,423(6941):762-769.
[9]Yamamoto Y,Hirose H,Saito I,etal.Correlation of the adipocyte-derived protein adiponectin with insulin resistance index and serum high-density lipoprotein-cholesterol,independent of body mass index,in the Japanese population[J]. Clin Sci,2002,103 (2):137-142.
[10]Tschritter O,Fritsche A,Thamer C,etal.Plasma adiponectin concentration predict insulin sensitivity of both glucose and lipid metabolism[J]. Diabetes,2003,52(2):239-243.
[11]Maeda K,Okubo K,Shimomura I,etal.cDNA cloning and expression of a novel adipose specific collagen-like factor,apM1 (Adipose Most abundant Gene transcript)[J]. Biochem Biophys Res Commun,1996,221(2):286-289.
[12]Maeda N,Shimomura I,Kishida K,etal.Diet-induced insulin resistance in mice lacking adiponectin/ACRP30[J]. Nat Med,2002,8:731-737.
[13]Yamauchi T,Kamon J,Minokoshi Y,etal.Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase[J]. Nat Med,2002,8:1288-1295.
[编辑] 一 凡
Q781
A
1673-1409(2009)02-R001-04
10.3969/j.issn.1673-1409(R).2009.02.001
2009-05-06
福建省重大科技专题项目(2008NZ0001-4);福建省自然科学基金项目(2009J01058);福建省科技项目(2008F5010);福建省科技创新平台项目(2007S1001)
刘峰(1976-), 男, 河南固始人,助理研究员,博士生,从事品质生物技术研究。
