外源褪黑素对盐旱复合胁迫下油莎豆种子萌发和幼苗生长的影响
2025-02-19王宁宁罗雪梅陈明媛郭睿刘建国



摘要:为探讨外源褪黑素对盐旱胁迫下油莎豆种子萌发和幼苗生长的缓解效应,采用0、50、100、150、200、250 μmol·L-1的褪黑素对油莎豆种子进行浸种引发处理,以200 mmol·L-1 NaCl溶液模拟盐胁迫、20% PEG-6000溶液模拟干旱胁迫,测定和分析不同处理下油莎豆种子的萌发、胚根和胚芽长度、生物量积累等指标。结果表明,200 mmol·L-1 NaCl胁迫、20% PEG-6000胁迫对油莎豆种子萌发和幼苗生长有显著抑制作用;50~150 μmol·L-1的褪黑素浸种对盐、旱胁迫下油莎豆种子的萌发和幼苗生长有促进作用,200~250 μmol·L-1的褪黑素浸种则抑制种子萌发和幼苗生长;盐旱复合胁迫加重了对油莎豆种子萌发和幼苗生长的危害程度,而外源褪黑素对油莎豆种子萌发指标、幼苗胚根和胚芽长度、植株干鲜重均有积极作用。综上所述,外源褪黑素对盐旱胁迫下油莎豆的种子萌发和幼苗生长有促进作用,其中150 μmol·L-1的褪黑素浸种对盐旱复合胁迫下种子萌发的缓解效果最佳,可有效减轻盐旱胁迫对种子萌发的负面效应。
关键词:褪黑素;盐旱复合胁迫;油莎豆;种子萌发;幼苗生长doi:10.13304/j.nykjdb.2023.0306
中图分类号:S565.9 文献标志码:A 文章编号:1008‐0864(2025)02‐0051‐11
油莎豆(Cyperus esculentus L.) 为莎草科(Cyperaceae)莎草属(Cyperus)的草本植物,是一种有较高综合利用价值的新型经济作物,具有粮、油、饲、药等多种用途。油莎豆具有适应性广、抗逆性强、生物量大、高附加值等优良特性,极具开发价值[1]。尤其油莎豆根系发达、分蘖强,生长2个月左右就基本可以覆盖地面,是天然的防风固沙材料[2]。新疆地处欧亚大陆腹地,属于典型的干旱半干旱区,水资源短缺、时空分布不均、蒸发量大[3]。土壤蒸发引起地下水沿土壤毛细管上移,同时土壤中的盐分也随着水分运动,从而形成土壤盐渍化[4]。随着干旱程度的增加,土壤盐渍化日益加重,干旱及土壤盐渍化已成为新疆农牧业发展最主要的自然灾害。因而,采取化学调控措施、提高植物抗逆能力是缓解土壤干旱及盐渍化对植物影响的有效生物措施。
褪黑素(melatonine,MT)化学名称为N- 乙酰-5-甲氧基色胺,是一种新型的植物生长调节剂,在植物生长发育、生理过程调控以及提高植物对生物和非生物胁迫响应等过程中具有重要作用。外源MT可有效减轻低温对菜豆种子萌发造成的渗透胁迫和氧化损伤程度,从而提高种子抵御低温的能力[5]; MT浸种可缓解盐胁迫对荞麦种子萌发及幼苗生长的抑制作用[6];外源MT能够有效缓解PEG造成的渗透调节胁迫对黄瓜种子萌发的抑制作用,并增强根系活力[7]。研究表明,MT作为抗氧化剂,能够通过提高非生物胁迫下抗氧化剂含量和抗氧化酶活性来清除过量的活性氧自由基,从而保护植物细胞免受氧化损伤,起到缓解逆境胁迫的作用[8]。目前,外源MT对干旱、盐碱胁迫下油莎豆种子萌发和幼苗生长的调控机理尚不明晰。因此,本研究分析外源施加褪黑素对盐旱胁迫下油莎豆种子萌发、生长的调控效应,旨在为新疆干旱区盐碱地开发利用以及促进盐碱地油莎豆的生产应用提供理论依据和技术参考。
1 材料与方法
1.1 试验材料
供试材料为‘中油莎1号’,将种子在试验前用5%次氯酸钠溶液处理10 min后用蒸馏水冲洗5次,35 ℃蒸馏水浸种48 h,滤纸吸干表面水分后自然风干。选取籽粒大小均一、饱满的种子用于发芽试验。试验所用NaCl、PEG-6000、褪黑素(MT)和无水乙醇均为分析纯。
1.2 试验设计
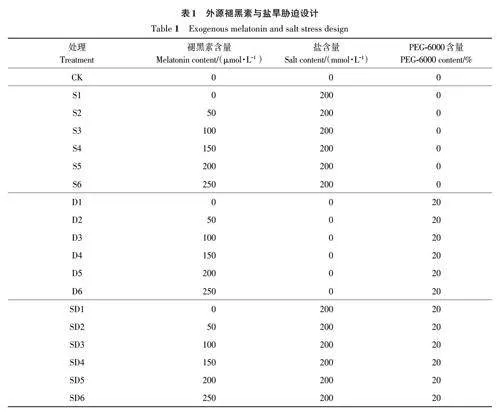
以200 mmol·L-1 NaCl进行盐(salt, S)胁迫处理、20% PEG-6000 进行干旱(drought,D)胁迫处理;将种子分别浸泡在0、50、100、150、200、250 μmol·L-1 的褪黑素溶液(MT)中35 ℃避光进行引发,浸种时间为24 h,共设置19个处理(表1),每个处理6次重复。将MT浸种后的种子用蒸馏水冲洗3次,随后将种子均匀置于装有蛭石的发芽盒中,每盒10粒种子,播深为5 cm,初始倒入350 mL培养液进行培养,盐胁迫以200 mmol·L-1 NaCl 溶液作为培养液,干旱胁迫20% PEG-6000溶液作为培养液,盐旱(salt and drought,SD)复合胁迫以200 mmol·L-1 NaCl+20% PEG-6000溶液作为培养液。置于35 ℃光温培养箱中,黑暗条件培养同化7 d后,转移至35 ℃的长日照(16 h光照,8 h黑暗)培养3 d,期间通过每天称重保持各培养皿含水量相同。
1.3 测定指标
1.3.1 种子萌发的测定 发芽试验共进行10 d,从第3天开始每天定时记录每个发芽盒中的发芽种子数。用第3 天的发芽种子数计算发芽势(germination potential,GP);第7 天统计发芽率(germination rate,GR);第10 天计算发芽指数(germination index,GI)、活力指数(vigor index,VI),公式如下。
1.3.2 植株生长指标的测定 播种后第10天选取有代表性的幼苗用蒸馏水冲洗干净,滤纸吸干水分测定胚芽长(germ length,GL)、胚根长(radicle length,RL)及植株鲜重(plant freshweight,P-FW);105 ℃杀青30 min后,80 ℃烘干至恒重测定植株干重(plant dry weight,P-DW)。
1.4 数据分析
利用Excel 2010 软件整理数据,SigmaPlot14.0 处理数据、绘制图表,SPSS 27.0软件进行显著性和相关性分析,采用 LSD(least significantdifference)法比较不同处理间的差异显著性。
2 结果与分析
2.1 褪黑素对盐胁迫下油莎豆种子萌发和幼苗生长的影响
2.1.1 对油莎豆种子萌发的影响 200 mmol·L-1NaCl胁迫下,油莎豆种子的发芽势、发芽率、发芽指数和活力指数均显著低于CK(图1)。与CK相比,S1处理油莎豆种子的发芽势、发芽率、发芽指数和活力指数分别降低52.00%、14.28%、21.53%和42.41%;随着褪黑素用量的增加,油莎豆种子的发芽势、发芽率、发芽指数和活力指数均呈先升高后降低趋势,且在50 μmol·L-1 褪黑素(S2)处理时达到最大值,分别为60%、90%、11.69 和161.64,显著高于S1处理(除发芽率外);S3处理下油莎豆种子仍有较高的发芽率,为76.67%;而S5处理下油莎豆种子的发芽势、发芽率、发芽指数和活力指数均显著降低,较S1 处理分别降低8.33%、8.34%、10.78%和10.55%。
2.1.2 对油莎豆幼苗生长的影响 盐胁迫对油莎豆幼苗生长有明显抑制作用(图2)。S1处理下油莎豆种子的胚芽长、胚根长分别为8.77、12.58 cm,较CK分别显著降低43.13%、26.39%;植株鲜重、植株干重分别为0.504 、0.071 g,较CK 分别显著降低65.31%和64.5%。由此表明,盐胁迫显著抑制油莎豆幼苗的生长。随着褪黑素用量的增加,盐胁迫下幼苗生长整体呈先增加后降低的趋势,其中S3处理的胚芽长和胚根长显著高于S1处理;S4 处理最高,胚芽长、胚根长分别为11.47、14.22 cm,较S1 相比分别显著增加30.79%、13.04%,植株鲜重、干重分别为0.867 和0.123 g,较S1分别显著增加72.02% 和73.24%;S2、S5、S6处理的胚芽长、胚根长及植株鲜重、干重与S1处理差异不显著。
2.2 褪黑素对干旱胁迫下油莎豆种子萌发和幼苗生长的影响
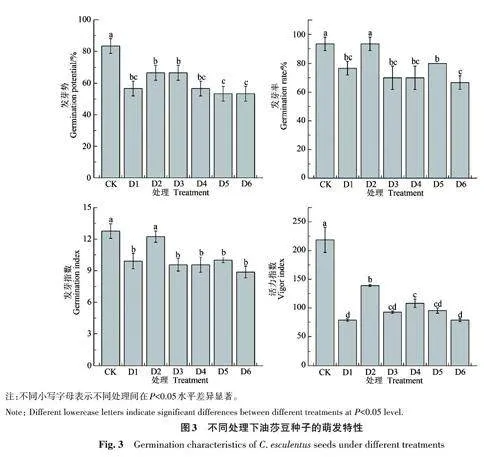
2.2.1 对油莎豆种子萌发的影响 不同褪黑素处理下,干旱胁迫试验中油莎豆种子各萌发指标的变化趋势与NaCl模拟盐胁迫试验下的变化趋势相似(图3)。相比于CK,D1处理种子的发芽势、发芽率、发芽指数和活力指数分别显著降低31.99%、17.85%、22.40% 和63.93%;D2 处理种子的发芽率、发芽指数和活力指数分别为93.33%、12.24和139.98,显著高于D1处理,且与CK 差异不显著(除活力指数外);从D3处理开始,随着褪黑素用量的增加,各萌发指标均呈现下降趋势,D6处理种子的发芽势、发芽率、发芽指数和活力指数最低,较D1分别降低5.89%、13.04%、10.49%和0.04%,但差异不显著。
2.2.2 对油莎豆幼苗生长的影响 由图4可知,干旱胁迫对油莎豆幼苗生长有明显抑制作用。CK处理的胚芽、胚根长度及植株鲜重、干重最高;D1处理的胚芽、胚根长度及植株鲜重、干重较CK分别显著降低51.43%、53.07%、78.46% 和76.00%,这表明干旱胁迫显著抑制了油莎豆胚芽、胚根的生长和生物量的积累。施加褪黑素后,油莎豆幼苗的生长随着褪黑素用量的增加呈先升高后降低趋势。D2、D5、D6处理与D1处理差异不显著;D3、D4处理的幼苗生长优于D1处理。其中D3 处理的胚根长和植株干重显著高于D1 处理;D4处理的胚芽长、胚根长及植株鲜重、干重较D1处理分别显著增加44.14%、41.27%、90.42%和60.42%。
2.3 褪黑素对盐旱复合胁迫下油莎豆种子萌发和幼苗生长的影响
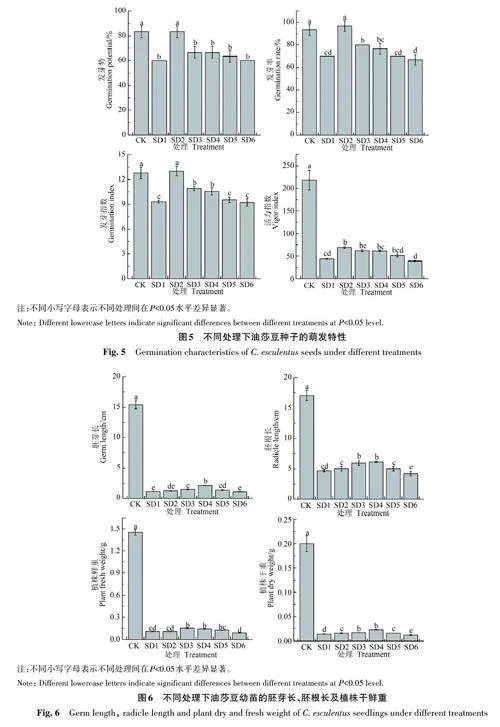
2.3.1 对油莎豆种子萌发的影响 由图5可知,随着褪黑素用量的增加,种子的萌发活力呈先升高后降低趋势,表明施用适宜量的褪黑素溶液对盐旱复合胁迫下油莎豆种子的萌发具有促进作用。与CK相比,SD1处理的发芽势、发芽率发芽指数和活力指数分别显著降低30.77%、25.00%、27.17%和80.07%,表明盐旱复合胁迫显著抑制了油莎豆种子的萌发。SD2处理种子的发芽势、发芽率、发芽指数和活力指数最高,分别为83.33%、96.67%、13.00和69.08,较SD1处理分别显著增加38.88%、38.10%、39.78%和58.44%;且发芽势、发芽率与CK差异不显著。从SD3处理开始,种子的萌发和活力逐渐降低,其中SD3和SD4处理的发芽指数显著高于SD1处理;SD5和SD6处理与SD1差异不显著。
2.3.2 对油莎豆幼苗生长的影响 由图6可知,SD1 处理的胚芽长、胚根长及植株鲜重、干重较CK 分别显著降低92.48%、72.56%、92.77% 和93.00%。随着褪黑素用量的增加,幼苗生长呈先升高后降低趋势。 SD2处理幼苗的生长与SD1处理差异不显著(除植株干重外);SD3和SD4处理幼苗的生长显著优于SD1处理,其中SD4处理最高,其胚芽长、胚根长分别为2.14、6.16 cm,较SD1分别显著增加84.48%、31.34%,植株鲜重、干重分别为0.14 和0.023 g,较SD1 处理分别显著增加33.33%、64.29%;SD6处理的胚根长、植株干重较SD1处理分别显著降低9.81%、14.29%。
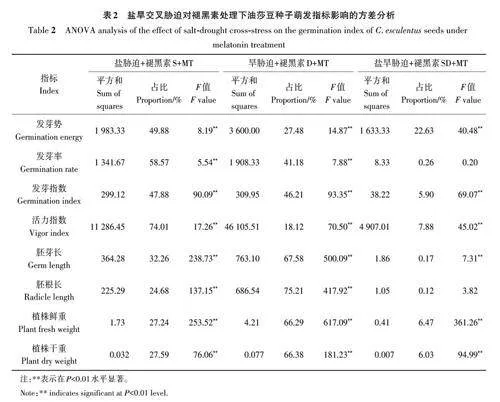
2.4 单一胁迫与复合胁迫对油莎生长影响差异的方差分析
方差分析(表2)表明,与盐胁迫和盐旱复合胁迫相比,干旱胁迫下施用褪黑素对油莎豆种子萌发、幼苗生长及生物积累量的影响较大。
2.5 油莎豆种子及幼苗耐盐旱能力评价
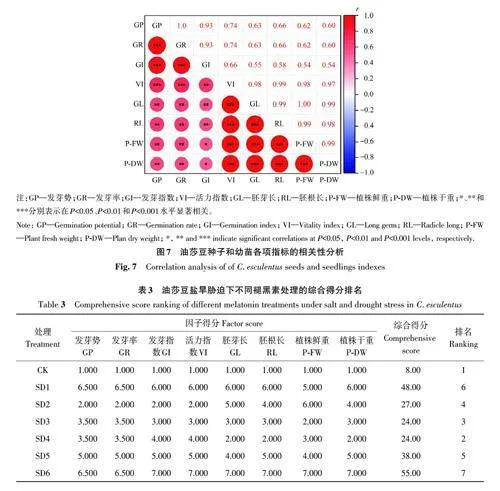
2.5.1 相关性分析 相关性分析(图7)表明,油莎豆种子的发芽率、发芽势、发芽指数、活力指数之间均存在极显著正相关关系;胚芽长、胚根长、植株鲜重、植株干重间也均存在极显著正相关关系,与发芽率之间呈显著正相关(Plt;0.05)。从指标间的相关性矩阵图中可以看出,各指标之间均有一定关联性,使其产生的生长效应发生重叠,因而对不同浓度褪黑素缓释效应的评价应该联系多个指标共同评估才具有可靠性。
2.5.2 综合性评价分析 为探讨不同水平褪黑素对盐旱胁迫下油莎豆影响的差异比较,采用个案排秩加法分别对不同水平褪黑素处理下的缓释作用进行综合性评价,结果(表3)表明,各处理对油莎豆种子萌发及幼苗生长的缓释效果表现为:CKgt;SD4gt;SD3gt;SD2gt;SD5gt;SD1gt;SD6,即150 μmol·L-1 褪黑素的缓释效应最佳。
3 讨论
种子萌发在植物的生长发育进程中起关键作用,是评判植物群体能否在逆境条件下顺利建植成功的决定性因素[9]。而苗期作为植物生长发育的关键时期,在植物的定植和繁衍过程中具有重要意义[10]。发芽率、发芽势、发芽指数和活力指数是反映种子发芽速度、发芽整齐度以及幼苗健壮程度的指标,常被用来评价种子的发芽活力,反映种子的萌发能力[11],可作为鉴定种子耐受能力的重要指标。生物积累量的变化作为植物对逆境胁迫响应的综合性表现,可作为评价作物耐盐抗旱能力最直观的指标[12]。
逆境胁迫条件下作物的外部形态及种子萌发率是作物遭受胁迫影响的最直接表现,受胁迫程度越大,种子萌发率越低,导致作物萌发出苗不齐,影响最终产量[13]。新疆作为典型的干旱半干旱地区,分布着大面积的盐渍土,而培育耐盐、抗旱的作物品种已然成为缓解新疆干旱盐渍化土壤对作物的胁迫影响及实现新疆农业可持续发展的重要任务。本研究发现,200 mmol·L-1 NaCl胁迫和20% PEG胁迫均会对油莎豆种子的发芽势、发芽率、发芽指数和活力指数有显著抑制作用,降低其生物累积量,与王志恒等[9]研究结果一致。这可能是因为盐、旱胁迫均会降低种子的吸水能力,从而导致种子内部水分亏缺,影响渗透调节,渗透压升高、水势降低,使得种子内部因吸水困难而产生生理干旱,进而极大程度地降低种子的发芽率、发芽势、发芽指数和活力指数,延长种子的萌发进程,严重时甚至会使种子失去萌发能力[14]。
为抵御外界不良生存环境对植物所造成的伤害,植物在长期的进化过程中逐渐演化出一系列复杂的防御机制[15]。褪黑素作为植物体内重要的吲哚类激素,其主要功能是作为抗氧化剂缓解植物的氧化损伤,作为一种生长调节剂,它能够有效提高植物对外界非生物胁迫的适应能力,增强植物的抗逆性[16]。Kolodziejczy等[17]用外源褪黑素处理玉米种子,能够提高种子的发芽率和芽鲜重。褪黑素的作用效果对剂量有很强的依赖性,适宜水平的褪黑素可以提高作物抵抗外界非生物胁迫的能力,但不同作物种类的最适水平可能存在差异[18]。董秋丽等[19]发现,施用适宜水平的外源褪黑素能够有效提高干旱胁迫下达乌里胡枝子种子的萌发活力;李平平等[20]研究表明,最适水平下的外源褪黑素能够通过促进幼苗形态生长,缓解盐碱胁迫对藜麦的生长抑制,进而增强藜麦的耐盐碱能力。
研究表明,对于同一植物而言,不同水平的褪黑素可以对胁迫下的植物起到正向缓释作用,缓解逆境胁迫对植物损伤,也可能会加剧植物受到的胁迫损伤[21]。李平等[22]研究发现,低水平褪黑素可以提高谷子在干旱胁迫下的萌发能力,胚芽和胚根的生长,提高谷子的生物量积累,增强谷子的耐旱性,而高水平褪黑素则会抑制谷子的萌发和生长。聂必林等[15]研究表明,中低水平的外源褪黑素对黑果枸杞种子萌发和幼苗生长有显著促进作用,而高水平则会产生抑制作用。本研究也得到了类似的结果,外源褪黑素对油莎豆种子的萌发和幼苗生长的影响表现为低水平下促进、高水平下抑制。褪黑素对油莎豆种子萌发、生物量的积累的影响表现为:随褪黑素水平的增加整体呈先升高后下降趋势;在褪黑素水平较低时对油莎豆种子萌发和幼苗生长影响较小,但中高水平下褪黑素影响较大,含量过高时甚至会抑制种子萌发和幼苗生长。
本研究发现,低浓度(50、100、150 μmol·L-1)下的外源褪黑素可促进盐、旱胁迫下油莎豆种子的萌发及幼苗生长,尤其是胚芽长度、植株鲜重。50 μmol·L-1外源褪黑素浸种可提高种子活力,从而有效提高非生物胁迫条件下种子的发芽势、发芽率、发芽指数和活力指数,使得盐旱胁迫下油莎豆种子有较高的发芽水平。这可能是因为褪黑素具有亲水和亲脂特性,容易进入到种子内部,且褪黑素作为抗氧化剂,可清除种子内的活性氧,保护种子免受氧化伤害,从而提高种子的活力和萌发率[23]。而150 μmol·L-1褪黑素对幼苗胚芽和胚根的生长及生物量积累的促进效果最为显著,这与李平平等[20]对盐碱胁迫下藜麦的研究结果相似。这可能是因为褪黑素在植物体内作为一种强抗氧化剂可清除植物体内产生的各种活性氧,提高植物体内抗氧化酶活性,从而保护植物免受氧化胁迫,且通过提高内源吲哚乙酸的含量促进根系生长[21,23‐24]。高浓度(200、250 μmol·L-1)外源褪黑素则会导致其促进油莎豆种子萌发和幼苗生长的作用不明显,甚至会抑制油莎豆的种子萌发和幼苗生长,这与张珂等[25]研究结果一致。这可能是因为高浓度褪黑素会与脱落酸、赤霉素之间呈现较强的协同作用,产生过强的抗氧化能力,但过强的抗氧化性能反向抑制了种子的萌发和幼苗的生长[26‐27]。此外,本研究通过对盐旱复合胁迫下油莎豆种子萌发指标、幼苗生长指标的方差分析结果可得,干旱胁迫下施用褪黑素是缓解种子萌发及幼苗生长的主要因子,在盐旱复合胁迫下,150 μmol·L-1褪黑素对作物的逆境胁迫的缓释效果最佳。
综上所述,褪黑素用于缓解盐旱胁迫对油莎豆种子萌发和幼苗生长的影响具有一定可行性,但应用于实际生产时则需要根据不同的生育进程合理施用,以缓解盐旱胁迫对油莎豆的胁迫损伤,提高油莎豆的耐盐抗旱性,从而有效提高盐旱胁迫下油莎豆种子的育苗成活率及幼苗的定植成功率,提高油莎豆的生物积累量,从而实现新疆干旱盐渍化地区油莎豆优质高产的目的。
参考文献
[1] 王瑞元,王晓松,相海,等.一种多用途的新兴油料作物——
油莎豆[J].中国油脂,2019,44(1):1-4.
[2] 阳振乐.油莎豆的特性及其研究进展[J].北方园艺,2017(17):
192-201.
YANG Z L. Characteristics and research progress of Cyperus
esculent [J]. Northern Hortic., 2017(17):192-201.
[3] 张喜成,徐长春,宋佳,等.基于遥感DSI的新疆干旱特征分
析[J].江苏农业科学,2020,48(2):239-246.
[4] 朱生堡,乌尔古丽·托尔逊,唐光木,等.新疆盐碱地变化及
其治理措施研究进展[J].山东农业科学,2023,55(3):158-165.
ZHU S B, Wuerguli Tuoerxun, TANG G M, et al .. Research
progress on saline-alkali land changes and its treatment measures
in Xinjiang [J]. Shandong Agric. Sci., 2023, 55(3):158-165.
[5] 杨晓旭,李梦娣,刘大军,等.外源褪黑素对低温胁迫下菜豆种
子萌发及抗性的影响[J].中国农学通报,2022,38(33):34-38.
YANG X X, LI M D, LIU D J, et al .. Exogenous melatonin
affects the germination and resistance ability of common bean
seeds under chilling stress [J]. Chin. Agric. Sci. Bull., 2022,
38(33):34-38.
[6] 雷新慧,万晨茜,陶金才,等.褪黑素与2,4-表油菜素内酯浸种
对盐胁迫下荞麦发芽与幼苗生长的促进效应[J].作物学报,
2022,48(5):1210-1221.
LEI X H, WAN C Q, TAO J C, et al .. Effects of soaking seeds
with MT and EBR on germination and seedling growth in
buckwheat under salt stress [J]. Acta Agron. Sin., 2022, 48(5):
1210-1221.
[7] ZHANG N, ZHAO B, ZHANG H J, et al .. Melatonin promotes
water-stress tolerance, lateral root formation, and seed
germination in cucumber (Cucumis sativus L.). [J]. J. Pineal
Res., 2013, 54(1):15-23.
[8] 王明瑶,曹亮,于奇,等.褪黑素浸种对盐碱胁迫下大豆种子
萌发的影响[J].作物杂志,2019(6):195-202.
WANG M Y, CAO L, YU Q, et al .. Effects of melatonin soaking
on germination of soybean seeds under saline-alkali stress [J].
Crops, 2019(6):195-202.
[9] 王志恒,杨秀柳,邹芳,等.旱盐交叉胁迫对甜高粱种子萌发
和生理特性的影响[J].中国农业科技导报,2021,23(2):37-49.
WANG Z H, YANG X L, ZOU F, et al .. Effects of salt and
drought cross stress on germination and physiological characteristics
of sweet sorghum seeds [J]. J. Agric. Sci. Technol., 2021, 23(2):
37-49.
[10] 塔伊尔·买买提江,马亚丽,兰海燕,等.盐旱耦合胁迫对费
尔干猪毛菜种子萌发及其幼苗生长的影响[J].干旱区研究,
2019,36(4):878-885.
Tayier Maimaitijiang, MA Y L, LAN H Y, et al .. Effects of salt
coupled with drought stress on seed germination and seedling
growth of Salsola ferganica [J]. Arid Zone Res., 2019, 36(4):
878-885.
[11] 余如刚,杜雪玲,陈楚,等.PEG胁迫对三种豆科牧草种子萌发
及幼苗生理影响[J].干旱地区农业研究,2012,30(5):99-103.
YU R G, DU X L, CHEN C, et al .. Effect of PEG stress on seed
germination and seeding physiology of three legumes [J]. Agric.
Res. Arid Areas, 2012, 30(5):99-103.
[12] 张睿,封晓辉,吴玉洁,等.长穗偃麦草(Thinopyrum ponticum)
幼苗对盐旱胁迫的生理响应[J].中国生态农业学报, 2022,
30(11):1795-1806.
ZHANG R, FENG X H, WU Y J, et al .. Interactive effects of
drought and salt stresses on the growth and physiological
characteristics of thinopyrum ponticum [J]. Chin. J. Eco-Agric.,
2022, 30(11):1795-1806.
[13] 姚海梅,李永生,张同祯,等.旱-盐复合胁迫对玉米种子萌发
和生理特性的影响[J].应用生态学报,2016,27(7):2301-2307.
YAO H M, LI Y S, ZHANG T Z, et al .. Effects of combined
drought and salinity stress on germination and physiological
characteristics of maize (Zea mays) [J]. Chin. J. Appl. Ecol.,
2016, 27(7):2301-2307.
[14] 梁龙飞,代胜,孙文涛,等. 盐旱交互胁迫对多花黑麦草
(Lolium multilorum)种子萌发及胚生长的影响[J].分子植物
育种,2020,18(10):3410-3420.
LIANG L F, DAI S, SUN W T, et al .. Effects of interactions
between Ca2+-salt and drought stress on seed germination and
embryo growth of Lolium multilorum [J]. Mol. Plant Breeding,
2020, 18(10):3410-3420.
[15] 聂必林,巫利梅,如马南木·尼合买提,等.外源褪黑素对复合
盐碱胁迫下黑果枸杞种子萌发和幼苗生长的缓解效应[J]. 西
北植物学报,2021,41(10):1711-1724.
NIE B L, WU L M, Rumananmu Nihemaiti, et al .. Alleviating
effects of exogenous melatonine on seed germination and seedling
growth of Lycium ruthenicum under complex salt-alkali stress [J].
Acta Bot. Bor-Occid. Sin., 2021, 41(10):1711-1724.
[16] MENENDEZ-PELAEZ A, POEGGELER B, REITER R J,
et al .. Nuclear localization of melatonin in different mammalian
tissues: immunocytochemical and radioimmunoassay evidence [J].
J. Cellular Biochem., 1993, 53(4):373-382.
[17] KOLODZIEJCZY K, IZABEL A, DZITK O, et al .. Exogenous
melatonin expediently modifies proteome of maize (Zea mays
L.) embryo during seed germination [J]. Acta Physiol. Plant,
2016, 38(6):1-18.
[18] 赵丽娟,麻冬梅,王文静,等.外源褪黑素对盐胁迫下紫花苜
蓿幼苗抗氧化能力以及光合作用效率的影响[J].西北植物
学报,2021,41(8):1355-1363.
ZHAO L J, MA D M, WANG W J, et al .. Effect of exogenous
melatonin on antioxidant capacity and photosynthetic efficiency of
alfalfa seedling under salt stress [J]. Acta Bot. Bor-Occid. Sin.,
2021, 41(8):1355-1363.
[19] 董秋丽,王聪聪,郑川,等.褪黑素引发对达乌里胡枝子种子
干旱萌发的影响[J].中国草地学报,2022,44(7):114-120.
DONG Q L, WANG C C, ZHEN C, et al .. Effects of melatonin
priming on germination of Lespedezad avurica seeds under
drought stress [J]. Chin. J. Grass., 2022, 44(7):114-120.
[20] 李平平,张永清,张萌,等.褪黑素浸种对混合盐碱胁迫下藜
麦生长及生理的影响[J].江苏农业学,2023,51(4):77-84.
[21] 陈贤,杨勇,刘凤权,等.植物褪黑素的研究进展[J].江苏农业
科学,2020,48(24):17-24.
[22] 李平,谢淑芹,方璐斌,等.干旱胁迫下不同褪黑素浓度处理
下谷子种子萌发特性[J].山西农业大学学报(自然科学版),
2022,42(5):76-83.
LI P, XIE S Q, FANG L B, et al .. Research on the impacts of
melatonin at different concentrations on the germination of
foxtail millet seeds under drought-stress conditions [J]. J.
Shanxi Agric. Univ. (Nat. Sci.), 2022, 42(5):76-83.
[23] 赵燕,王东华,赵曦阳,等.植物中褪黑素的研究进展[J].西北
植物学报,2014,34(1):196-205.
ZHAO Y, WANG D H, ZHAO X Y, et al .. Recent advance on
the melatonin in plant [J]. Acta Bot. Bor-Occid. Sin., 2014,
34(1):196-205.
[24] 李波,郝瑞婷,孙晶,等.植物褪黑素研究进展及未来展望[J].
运城学院学报,2021,39(6):1-12.
LI B, HAO R T, SUN J, et al .. Research progress and future
prospects of plant melatonin [J]. J. Yuncheng Univ., 2021,
39(6):1-12.
[25] 张珂,李依,厉萌萌,等.外源褪黑素对镉胁迫下小麦幼苗生
长生理特征及镉含量的影响[J]. 轻工学报,2022,37(4):
111-117.
ZHANG K, LI Y, LI M M, et al .. Effects of exogenous
melatonin on growth and physiological characteristics and Cd
content of wheat seedling under Cd stresses [J]. J. Light Ind.,
2022, 37(4):111-117.
[26] LYU Y, PAN J J, WANG H P, et al .. Melatonin inhibits seed
germination by crosstalk with abscisic acid, gibberellin, and
auxin in Arabidopsis [J/OL]. J. Pineal Res., 2021, 70(4):
e12736 [2023-03-15]. https://doi.org/10.1111/jpi.12736.
[27] REITER R J, MAYO J C, TAN D X, et al .. Melatonin as an
antioxidant: under promises but over delivers [J]. J. Pineal
Res., 2016, 61(3):253-278.
