丁氏稻种萌发期耐热性筛选与评价
2025-01-28钟伟杰陈洁刘辉朱振权阮阳马帅鹏刘向东刘羽佳李翔








摘要" 本研究对P0340、CII141和R289等20份丁氏稻种在种子萌发期进行高温胁迫处理,以正常培养为对照,测定其发芽势、发芽率和根、芽长等10项生长指标,利用隶属函数综合评价法和分级评价值进行耐热综合评估,同时进行相关分析、回归分析及聚类分析,并结合过氧化氢(H2O2)和丙二醛(MDA)含量测定和超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)活性分析,探讨不同稻种萌发期生长指标和生理指标的差异,以筛选耐热性强的稻种。结果表明,高温处理后,水稻的发芽势等生长指标在品种间和处理间差异较大;耐热综合评估方面,P0340排位较高,耐热性强;Y595排位较低,耐热性弱;相关性分析和回归分析确立了发芽率和幼苗干重可作为耐热性鉴定的重要指标;聚类分析将参试稻种耐热性划分为极强、强、中等、弱和极弱5类,其中耐热性极强品种为P0340,耐热性强品种为CII141、R289和Y1544,耐热性中等品种包括R107和Δ133等10个,耐热性弱品种包括PII140和PII21等5个,耐热性极弱品种为Y595;生理指标方面,与对照相比,高温胁迫下P0340的H2O2和MDA的含量增幅较少,差异无统计学意义(Pgt;0.05);SOD、CAT和POD活性与对照相比显著增加(Plt;0.05),表明P0340萌发期耐热性强,活性氧含量累积少,抗氧化能力强。综上,筛选得到的高耐热稻种P0340,其可为培育耐热新品种提供材料基础。
关键词" 丁氏稻种;耐热性;萌发期;综合评价;生理特性
中图分类号" S326" " " "文献标识码" A" " " "文章编号" 1007-7731(2025)02-0001-09
DOI号" 10.16377/j.cnki.issn1007-7731.2025.02.001
Screening and evaluation of heat tolerance during the germination stage in Ting’s rice
ZHONG Weijie1,2" " CHEN Jie1,2" " LIU Hui2" " ZHU Zhenquan2" " RUAN Yang2" " MA Shuaipeng3" " LIU Xiangdong4
LIU Yujia1,2" " LI Xiang1,2
(1Guangdong Provincial Key Laboratory of Utilization and Conservation of Food and Medicinal Resources in Northern Region, Shaoguan University, Shaoguan 512005, China;
2College of Biology and Agriculture, Shaoguan University, Shaoguan 512005, China;
3Guangzhou Experimental Station of Chinese Academy of Tropical Agricultural Sciences, Guangzhou 510140, China;
4State Key Laboratory for Conservation and Utilization of Subtropical Agro-Bioresources, South China Agricultural
University, Guangzhou 510642, China;
Abstract" The study subjected 20 Ting’s rice varieties, including P0340, CII141, and R289 were treated with high temperature stress during seed germination,normal culture was used as control, 10 key germination indexes such as germination potential, germination rate, and root and shoot length were measured. By using the membership function comprehensive evaluation method and grading evaluation value, the heat resistance comprehensive evaluation was carried out. Meanwhile, correlation analysis, regression analysis and cluster analysis were carried out, combined with the determination of hydrogen peroxide (H2O2) and malondialdehyde (MDA) and the activity analysis of superoxide dismutase (SOD), catalase (CAT) and peroxidase (POD), the differences of growth indexes and physiological indexes of different rice varieties at germination stage were discussed, so as to screen rice varieties with strong heat resistance.The results showed that after high temperature treatment, the growth indexes such as germination potential of rice were different between varieties and treatments.In terms of heat resistance comprehensive evaluation, P0340 ranked first, showcasing strong heat tolerance, while Y595 ranked last, indicating weak heat tolerance. Correlation analysis and regression analysis established that germination rate and seedling dry weight could be used as important indexes for heat resistance identification.According to the cluster analysis, the heat resistance of the rice varieties tested was divided into 5 categories:extremely strong, strong, medium, weak and extremely weak. Among them, the variety with strong heat resistance was P0340, the varieties with strong heat resistance were CII141, R289, and Y1544, 10 varieties with medium heat resistance included R107 and Δ133, etc., and 5 varieties with weak heat resistance included PII140 and PII21,etc. The very weak heat resistance variety was Y595. In terms of physiological indexes, compared with control, under high temperature stress, H2O2 and MDA contents of P0340 increased less, the difference was not statistically significant (Pgt;0.05). The activities of SOD, CAT, and POD in P0340 significantly increased compared to the control (Plt;0.05). These findings suggest that P0340 displays strong heat tolerance during the germination stage, characterized by minimal accumulation of reactive oxygen species and enhanced antioxidant capacity. In conclusion, the identification of the high heat tolerance rice variety P0340 in this study offers a valuable genetic resource for the development of new heat tolerance varieties.
Keywords" Ting’s rice; heat tolerance;germination stage; comprehensive evaluation; physiological traits
水稻是主要的粮食作物之一,幼苗的萌发能力及早期生长状况是衡量其活力的重要指标。温度对其生长与分布至关重要,Zhao等[1]研究表明,每上升1 ℃,水稻的产量将下降2%。持续高温会降低水稻种子的发芽潜力,导致发芽率降低,幼苗活力减弱[2-3]。水稻幼苗期生长的适宜温度在25~28 ℃,在42~45 ℃的高温环境下,幼苗会出现严重的水分流失现象,叶片逐渐枯萎变黄,幼苗和根系的生长受到严重抑制,甚至导致幼苗死亡[4-6]。由于遗传背景不同,稻种对热胁迫的抗性存在显著差异。在45 ℃热胁迫条件下,粳稻日本晴的幼苗几乎全部枯萎,其耐热能力较弱[5];而籼稻HT54的幼苗则展现出较高的耐热性,能够耐受48 ℃的高温,持续79 h存活[7]。
筛选耐热稻种是解决水稻耐热性栽培的有效途径之一。近年来,研究人员针对不同稻种的幼苗期耐热性进行鉴定评价,并筛选获得耐热种质[8-10]。如刘进等[8]利用籼稻、粳稻核心种质资源,以耐热等级和幼苗存活率为指标,筛选出嘉育253、中优早8号和秀水09等20个耐热品种。季平等[9]认为水稻苗期叶片抗氧化酶活性和质膜损伤程度等可作为耐热水稻品种筛选的参考指标。稻种资源作为水稻育种工作中不可或缺的物质材料,承载着丰富的遗传信息。其中,丁氏稻种资源(Ting’s rice germplasm collection),以其独特性和多样性而备受瞩目[11]。该资源主要由3部分构成:一是丁颖院士收集的7 000余份各地栽培稻,二是卢永根院士收集的2 000余份野生稻资源,三是新型四倍体水稻新种质和水稻染色体单片段代换系。该稻种资源具有丰富的遗传变异性,含有优良的抗逆基因(耐冷、耐铝和耐锌等),为水稻育种提供了宝贵的材料[12-15]。目前,已利用丁氏稻种资源鉴定了一批优良的抗逆材料,但有关丁氏稻种萌发期耐热性的研究较少。因此,本研究利用广东省植物分子育种重点实验室构建的丁氏稻种核心资源,选取P0340和CII141等20种遗传多样性丰富的稻种作为研究材料,利用隶属函数综合评价等方法对稻种进行耐热性综合评价,测定过氧化氢(Hydrogen peroxide,H2O2)、丙二醛(Malondialdehyde,MDA)含量和超氧化物歧化酶(Superoxide dismutase,SOD)、过氧化氢酶(Catalase,CAT)及过氧化物酶(Peroxidase,POD)活性,探究高温胁迫对水稻苗期生长发育和生理代谢的影响,为耐热水稻品种的培育提供参考。
1 材料与方法
1.1 供试材料
选取20份丁氏稻种,分别为P0340、CII141、R289、Y1544、Δ134、Δ125、Δ133、R107、Δ151、Y1472-2、P0081、P0116、P0328、DR9、ZB132、Y814、PII140、PII21、P616和Y595,由华南农业大学亚热带农业生物资源保护与利用国家重点实验室提供。
1.2 试验试剂
生理指标测定试剂盒均采购于南京建成生物工程研究所,包括H2O2测定试剂盒(A064-1-1)、MDA测定试剂盒(A003-3-1)、SOD测试盒(A001-1-2)、CAT测定试剂盒(A007-1-1)和POD测定试剂盒(A084-3-1)。
1.3 试验方法
试验于2022—2023年在广东韶关学院生物与农业学院分子生物学与基因工程研究室进行。参照叶婵娟等[10]研究方法进行水稻种子的处理,每份水稻品种挑选200余粒饱满、大小一致的种子,使用10%次氯酸钠消毒30 min,消毒后用蒸馏水冲洗3次,然后加入蒸馏水至完全没过种子,在28 ℃恒温恒湿培养箱(暗培养)中浸种24 h;吸水后的种子转移到垫有两层湿润滤纸的培养皿(直径10 cm)中,在28 ℃恒温恒湿培养箱(暗培养)中进行催芽。种子露白后点播到96孔培养板中,用木村B水稻营养液(NS1050)进行培养,定期更换营养液,并置于人工气候箱中进行苗期耐热性处理。正常培养环境(对照)设置:28 ℃,13 L∶11 D,湿度80%。模拟高温环境设置:42 ℃,13 L∶11 D,湿度80%。高温胁迫处理7 d后测定幼苗生长指标,并将采集的幼苗进行液氮速冻,放入-80 ℃冰箱保存,用于后续生理指标测定。试验共设3次重复。
1.4 测定指标和方法
1.4.1 生长指标 参照Wang等[16]方法分别计算发芽势、发芽率、发芽指数和活力指数,计算如式(1)~(4)。测量第7天已发芽种子的芽长、根长和根数,以及幼苗鲜重。随后,将幼苗放入烘箱中烘干至恒重,称量幼苗干重,幼苗相对含水量计算如式(5)。
发芽势=第4天发芽的种子数/供试种子数 (1)
发芽率(%)=第7天发芽的种子数/供试种子数 (2)
发芽指数(%)=∑(每天发芽种子数/相应发芽天数)×100 (3)
活力指数=发芽指数×(第7天的芽长+第7天的根长) (4)
幼苗相对含水量(%)=(幼苗鲜重-幼苗干重)/幼苗鲜重 (5)
1.4.2 耐热系数 为去除各性状的单位差异,参照张娟伟等[17]方法,分别计算发芽势、发芽率、发芽指数、活力指数、芽长、根长、根数、幼苗鲜重、幼苗干重和幼苗相对含水量的耐热系数,计算如式(6)。
耐热系数=处理组指标测定值/对照指标测定值 (6)
1.4.3 耐热性综合评价 耐热性综合评价是将测量的各个单项指标(耐热系数)通过转化成新的综合指标,并采用加权平均法将各品种的隶属值累加,从而得出耐热综合评价值D值。D值越大,说明该品种的耐热性等级越高[18-19]。利用SPSS 21.0软件对发芽势、发芽率和发芽指数等10项指标的耐热系数进行主成分分析,采用最大方差法提取主成分,将多项指标转化为少数几个综合指标,其中累积贡献率大于85%,且特征数大于1,并根据各综合指标的主成分值计算隶属函数值、权重系数和耐热综合评价值,计算如式(7)~(9)。
μ(X_j)=(X_j-X_min)/(X_max-X_min) (7)
W_j=P_j/(∑P_j) (8)
D=∑[μ(X_j)×W_j] (9)
式(7)~(9)中,Xj表示第j个综合指标的主成分值(j=1,2,3,…,n),Xmax、Xmin分别为在所有稻种中第j个综合指标的最大值和最小值。Wj为权重,指第j个综合指标(主成分)在所有综合指标(主成分)中的重要程度;Pj指通过主成分分析所获得的第j个综合指标(主成分)的贡献率(j=1, 2, 3, …, n)。D值表示综合评价值(j=1, 2, 3, …, n)。
1.4.4 耐热分级评价值 参照文献[20]的划级方法计算耐热分级评价值GC,GC值越大,综合耐热性越强。以稻种的各耐热系数平均值x ̅_j为基准(j=1, 2, 3, …, n),以标准差s_j作为等级划分的依据。各稻种的耐热系数x_ij表示第i个稻种的第j个耐热系数(i=1, 2, 3, …, n;j=1, 2, 3, …, n),如:x_ij≥x ̅_j+s_j,则赋级别值1;x ̅_j+s_jgt;x_ij≥x ̅_j,则赋值2;x ̅_jgt;x_ij≥x ̅_j-s_j,则赋值3;x ̅_j-s_jgt;x_ij,则赋值4。将同一参试稻种各耐热系数下的级别值相加,得到各稻种的耐热级别值总和GV,耐热分级评价值GC计算如式(10)。
GC=(nm-GV)/(nm-n) (10)
式中,n为指标个数(本研究为10),m为分级级数(本研究为4)。
1.4.5 相关性分析、回归分析及聚类分析 利用SPSS 21.0软件对10项指标的耐热系数进行Pearson相关性分析。以各耐热系数为自变量,D值为因变量进行逐步分析,同时方差扩大因子值VIF少于5,建立多元回归方程。根据D值,采用平方欧式距离和组间平均连接法对20份稻种进行聚类分析。
1.4.6 生理指标 根据耐热综合评价值和聚类分析结果,挑选5份不同等级耐热能力的稻种进行生理指标测定,包括H2O2和MDA含量,SOD、CAT和POD活性。依据试剂盒说明书步骤进行各指标测定和计算。生理指标的相对含量或活性计算如式(11)。
相对含量或活性=(处理-对照)/对照 (11)
1.5 数据处理
采用Excel 2019软件对数据进行整理,使用SPSS 21.0软件进行主成分、相关性、回归及聚类分析,使用GraphPad Prism 9.5软件对数据进行差异分析及作图。
2 结果与分析
2.1 高温胁迫对水稻生长指标的影响
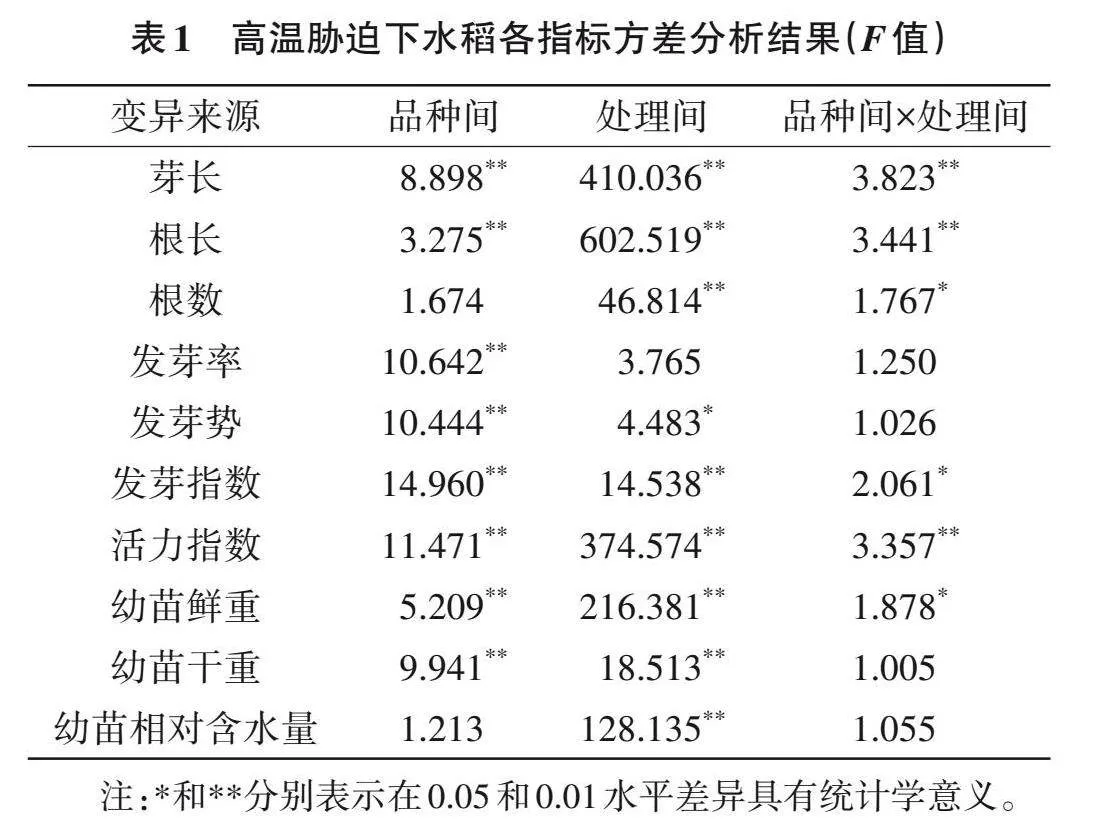
高温胁迫下,20个参试稻种的表型产生了变化,其中P0340和Y595最为明显(图1)。对10个生长指标进行测定,方差分析结果(F值)表明,根数和幼苗相对含水量在品种间差异无统计学意义(Pgt;0.05),芽长、根长、发芽率、发芽势、发芽指数、活力指数、幼苗鲜重和幼苗干重在品种间差异存在统计学意义(Plt;0.01);发芽率在处理间差异无统计学意义(Pgt;0.05),芽长、根长、根数、发芽势、发芽指数、活力指数、幼苗鲜重、幼苗干重和幼苗相对含水量在处理间差异存在统计学意义(Plt;0.05)(表1)。说明高温处理后,不同稻种的单项指标存在显著差异,直接利用单项指标不能准确评价稻种的耐热性。
2.2 耐热性综合评价及分级评价
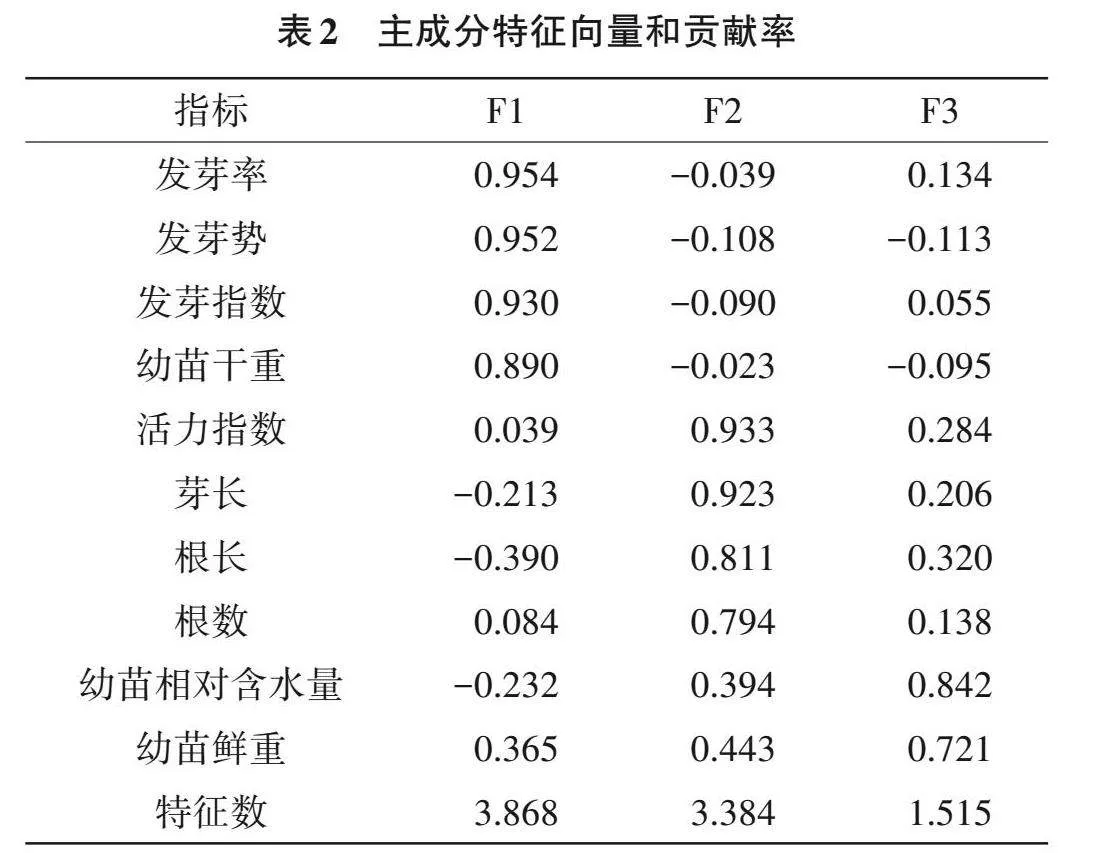
主成分分析提取了累积贡献率大于85%且特征值大于1的前3个主成分(F1,F2和F3),分别解释了38.681%、33.836%和15.151%的数据变异,合计可解释表型总变异的87.668%,可较好代表10个原始指标,即综合指标1、2和3。其中,发芽率、发芽势、发芽指数和幼苗干重包含在F1中,F2包含活力指数、芽长、根长和根数,F3包括幼苗相对含水量和幼苗鲜重(表2)。
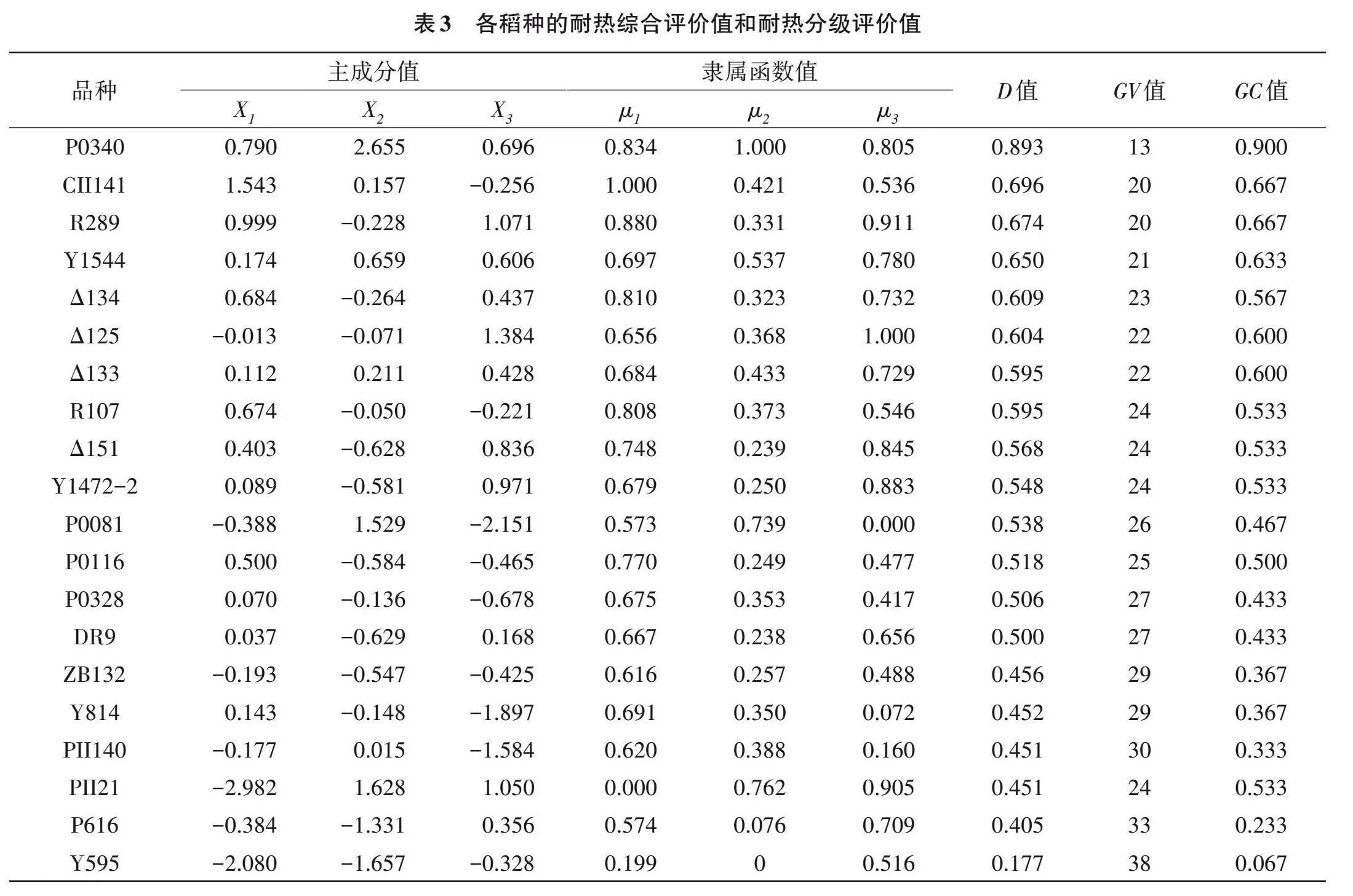
3个主成分的权重分别为44.122%、38.596%和17.282%(表2),主成分上的隶属函数值和耐热综合评价值结果显示(表3),稻种P0340 D值最高,为0.893,而Y595 D值最低,为0.177。此外,分级评价法结果显示,P0340的GC值最高,为0.900,而Y595的GC值最低,为0.067。综上,分级评价结果与隶属函数的综合评价法得出的结论相吻合,表明P0340耐热性较强;Y595耐热性较弱。
2.3 生长指标的相关性及回归分析
对萌发期的10个指标进行相关性分析,结果表明,发芽势与发芽指数、发芽率呈正相关(Plt;0.01),相关系数分别为0.93和0.90,活力指数与根长呈正相关(Plt;0.01),相关系数为0.87(图2)。相关系数较高意味着变量间存在信息的冗余现象,因此,在后续的多元回归分析中,为了减少变量间的干扰,移除发芽势、发芽指数和活力指数这3个变量。以D值为因变量,剩余7个指标为自变量进行逐步回归分析,并建立最优D值的线性回归模型:D=-0.903+0.611×发芽率+0.311×芽长+0.09×根数+0.441×幼苗干重+0.367×幼苗相对含水量(R2=0.97,Durbin-Watson=2.525,Plt;0.01,VIFlt;5)。该模型拟合度高(R2=0.97,接近1,Plt;0.01),各变量间不存在多重共线性(VIFlt;5),且误差项间不存在序列相关(Durbin-Watson接近中值2),符合建模前提,该模型有效且有价值。从模型系数来看,发芽率和幼苗干重对高温胁迫反应较为敏感,可作为耐热性鉴定的重要指标,其值越高,D值越高,表明稻种耐热性能越强。
2.4 参试稻种耐热能力的聚类分析
聚类分析可将20份稻种耐热性划分为极强、强、中等、弱和极弱5类(图3)。第Ⅰ类群为耐热性中等品种,包括R107、Δ133、Δ125、Δ134、DR9、P0328、P0116、Y1472-2、P0081和Δ151,共10个,占比50%;第Ⅱ类群为耐热性强品种,为CII141、R289和Y1544,占比15%;第Ⅲ类群为耐热性弱品种,包括PII140、PII21、Y814、ZB132和P616,共5个,占比25%;第Ⅳ类群为耐热性极强品种,为P0340;第Ⅴ类群为耐热性极弱品种,为Y595。
横轴数值代表平方欧式距离。
2.5 高温胁迫对水稻生理指标的影响
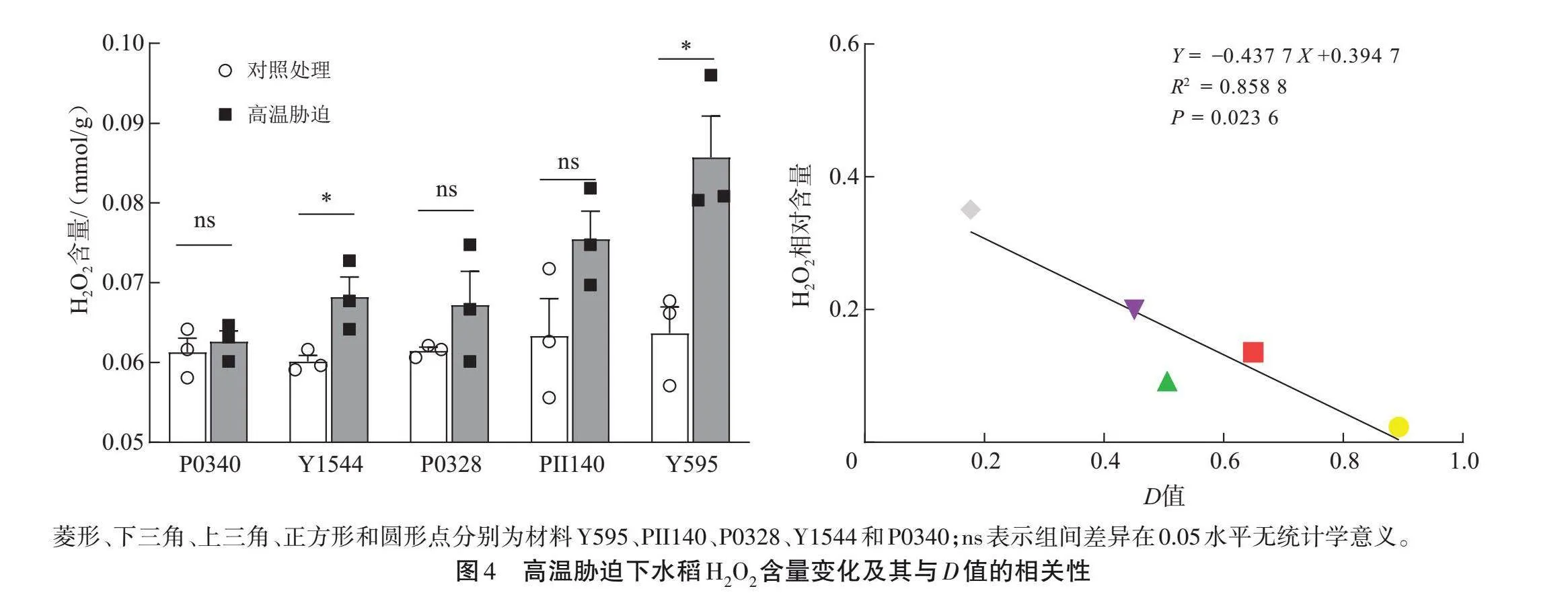
挑选P0340(极强)、Y1544(强)、P0328(中等)、PII140(弱)和Y595(极弱)5个稻种,测定H2O2和MDA含量,SOD、CAT和POD活性(图4~5)。Y595幼苗的H2O2和MDA含量较对照显著增加(Plt;0.05),而P0340幼苗的H2O2和MDA含量与对照组间的差异无统计学意义(Pgt;0.05);相关分析发现,H2O2和MDA相对含量与D值之间呈显著负相关(Plt;0.05),相关系数R2分别为0.858 8和0.831 0。说明不同耐热性稻种的H2O2和MDA含量存在差异,耐热性强的稻种在高温胁迫下MDA和H2O2含量变化较小。
SOD、CAT和POD的活性变化显示(图6),P0340幼苗的SOD、CAT和POD活性在高温处理后显著增加,而其他稻种的增加幅度不明显;相关分析发现,各稻种的SOD、CAT和POD相对活性与D值呈正相关,其中POD和CAT相对活性与D值之间的相关性差异存在统计学意义(Plt;0.05)。说明耐热性较强的稻种在高温胁迫下CAT和POD活性增幅较大。
菱形、下三角、上三角、正方形和圆形点分别为材料Y595、PII140、P0328、Y1544和P0340;ns表示组间差异在0.05水平无统计学意义。
3 结论与讨论
丁氏稻种资源丰富,其蕴含优良的抗逆基因,为水稻育种和种质创新提供了宝贵的物质材料[11]。本研究利用20份丁氏核心稻种,在其种子萌发期进行了高温胁迫处理,发现品种间对高温胁迫的响应存在明显差异。采用综合指标评价水稻耐热性的方法,相较于单一指标评估,更能全面、准确地反映水稻的耐热潜力,有效避免了评估的局限性。Zhang等[18]采用隶属函数法对114份稻种进行耐盐性筛选,得出耐盐性较强的稻种日光。李其勇等[20]研究表明,分级评价法和隶属函数法在稻种芽期耐旱性分析中展现出良好的鉴定能力。Yao等[21]基于该方法对6个水稻品种灌浆期的耐高温性进行综合评价,发现稻种黄华占耐热能力最好。本研究利用隶属函数法和分级评价法,对20份丁氏稻种进行耐热性评价,发现两种方法的评价结果高度一致,相互验证了P0340稻种在耐热性方面的优异表现。同时,本研究还构建了D值回归方程:D=-0.903+0.611×发芽率+0.311×芽长+0.09×根数+0.441×幼苗干重+0.367×幼苗相对含水量,发现发芽率和幼苗干重在高温胁迫下表现出极高的敏感性,即发芽率越高、幼苗干重越大,稻种的耐热综合评分值也相应越高。这2个指标不仅能够反映种子的萌发状态,还能有效反映稻种在萌发期的有效物质积累情况,为快速筛选和初步评估耐热性稻种提供参考。
持续的高温胁迫会导致活性氧(Reactive oxygen species,ROS)在细胞内过量积累,从而打破ROS稳态,引发氧化损伤,使水稻表现出生长迟缓、稻米白化及幼苗死亡等现象[22]。Zhao等[23]研究表明,高温胁迫会损害抗氧化酶活性,尤其是SOD和CAT,其在植物体内具有清除ROS、维持细胞稳态等作用。本研究发现,在高温胁迫下,候选稻种P0340的SOD、CAT和POD活性相较于对照显著升高,且增长幅度较为显著,表明稻种P0340在遭遇高温胁迫时,可通过增强这些抗氧化酶的活性,提升其耐热性。过量的ROS积累会进一步加剧膜脂过氧化和蛋白质氧化,损害生物膜的结构和功能,并导致过氧化物含量的增加。Bahuguna等[24]研究表明,稻种NERICA-L-44在高温胁迫下具有较高的抗氧化酶活性和较低的过氧化物含量,在水稻营养和生殖阶段表现出突出的耐热能力,可作为耐热性育种的新材料。本研究发现,候选稻种P0340在高温胁迫下,其MDA和H₂O₂的含量在胁迫处理前后并未发生显著变化,而稻种Y595的MDA和H₂O₂含量在高温胁迫下显著增加,说明稻种P0340在幼苗期表现出较好的耐热能力。本研究不仅证实了稻种P0340在高温胁迫下的耐热性,还进一步验证了利用耐热综合评价值D值来鉴定稻种萌发期耐热能力的可靠性。
综上,本研究利用综合评价方法对20 份丁氏稻种资源进行耐热性评价,通过耐热综合评价筛选出耐热性强的水稻品种P0340,并通过活性氧含量和抗氧化能力分析进一步验证了其耐热能力。在后续的研究中,还需要进一步考证P0340在大田中的具体表现。此外,P0340可作为候选耐热品种,与耐热性极弱品种Y595进行杂交,构建F2代分离群体,从而对耐热品种中携带的耐热基因进行初步定位。另外,还可以将P0340与当前主导稻种进行回交,实现对主导稻种耐热性的遗传改良,培育新型耐热稻种。
参考文献
[1] ZHAO C,LIU B,PIAO S L,et al. Temperature increase reduces global yields of major crops in four independent estimates[J]. Proceedings of the National Academy of Sciences of the United States of America,2017,114(35):9326-9331.
[2] FAHAD S,BAJWA A A,NAZIR U,et al. Crop production under drought and heat stress:plant responses and management options[J]. Frontiers in plant science,2017,8:1147.
[3] LIU J,HASANUZZAMAN M,WEN H L,et al. High temperature and drought stress cause abscisic acid and reactive oxygen species accumulation and suppress seed germination growth in rice[J]. Protoplasma,2019,256(5):1217-1227.
[4] LIU J P,SUN X J,XU F Y,et al. Suppression of OsMDHAR4 enhances heat tolerance by mediating H2O2-induced stomatal closure in rice plants[J]. Rice,2018,11(1):38.
[5] LIU J P,ZHANG C C,WEI C C,et al. The RING finger ubiquitin E3 ligase OsHTAS enhances heat tolerance by promoting H2O2-induced stomatal closure in rice[J]. Plant physiology,2015,170(1):429-443.
[6] KILASI N L,SINGH J,VALLEJOS C E,et al. Heat stress tolerance in rice (Oryza sativa L.):identification of quantitative trait loci and candidate genes for seedling growth under heat stress[J]. Frontiers in plant science,2018,9:1578.
[7] WEI H,LIU J P,WANG Y,et al. A dominant major locus in chromosome 9 of rice (Oryza sativa L.) confers tolerance to 48 ℃ high temperature at seedling stage.[J].The journal of heredity,2013,104(2):287-94.
[8] 刘进,崔迪,余丽琴,等. 水稻苗期耐热种质资源筛选及QTL定位[J]. 中国水稻科学,2022,36(3):259-268.
[9] 季平,刘经威,王欣婷,等. 不同水稻品种苗期叶片生理特性对高温胁迫的响应[J]. 西北农林科技大学学报(自然科学版),2024,52(1):36-48.
[10] 叶婵娟,陈可,周新桥,等. 4个籼稻品种苗期耐热性评价及分子机理分析[J]. 华南农业大学学报,2023,44(6):906-914.
[11] 陈志雄,王兰,吴锦文,等. 丁氏稻种资源有利基因挖掘与创新研究进展[J]. 华南农业大学学报,2023,44(5):649-658.
[12] SONG J Y,LI J Q,SUN J,et al. Genome-wide association mapping for cold tolerance in a core collection of rice (Oryza sativa L.) landraces by using high-density single nucleotide polymorphism markers from specific-locus amplified fragment sequencing[J]. Frontiers in plant science,2018,9:875.
[13] ZHANG P,ZHONG K Z,ZHONG Z Z,et al. Mining candidate gene for rice aluminum tolerance through genome wide association study and transcriptomic analysis[J]. BMC plant biology,2019,19(1):490.
[14] ZHONG K Z,XIE L H,HU S K,et al. Genome-wide association study of zinc toxicity tolerance within a rice core collection (Oryza sativa L.)[J]. Plants,2022,11(22):3138.
[15] GHOURI F,SHAHID M J,LIU J W,et al. Polyploidy and zinc oxide nanoparticles alleviated Cd toxicity in rice by modulating oxidative stress and expression levels of sucrose and metal-transporter genes[J]. Journal of hazardous materials,2023,448:130991.
[16] WANG Z F,WANG J F,BAO Y M,et al. Quantitative trait loci analysis for rice seed vigor during the germination stage[J]. Journal of Zhejiang university Science B,2010,11(12):958-964.
[17] 张娟伟,石亚飞,路旭平,等. 种子萌发期粳稻种质资源耐旱性综合评价[J]. 核农学报,2022,36(11):2093-2103.
[18] ZHANG R,HUSSAIN S,WANG Y,et al. Comprehensive evaluation of salt tolerance in rice (Oryza sativa L.) germplasm at the germination stage[J]. Agronomy,2021,11(8):1569.
[19] 戴海芳,武辉,阿曼古丽·买买提阿力,等. 不同基因型棉花苗期耐盐性分析及其鉴定指标筛选[J]. 中国农业科学,2014,47(7):1290-1300.
[20] 李其勇,朱从桦,李星月,等. 水稻芽期抗旱性综合评价及鉴定指标鉴选[J]. 西北农业学报,2023,32(1):18-32.
[21] YAO D P,WU J,LUO Q H,et al. Comprehensive evaluation of high temperature tolerance of six rice varieties during grain-filling period based on key starch physicochemical indexes[J]. LWT,2021,148:111804.
[22] XU Y F,CHU C C,YAO S G. The impact of high-temperature stress on rice:challenges and solutions[J]. The crop journal,2021,9(5):963-976.
[23] ZHAO Q,ZHOU L J,LIU J C,et al. Involvement of CAT in the detoxification of HT-induced ROS burst in rice anther and its relation to pollen fertility[J]. Plant cell reports,2018,37(5):741-757.
[24] BAHUGUNA R N,JHA J,PAL M,et al. Physiological and biochemical characterization of NERICA-L-44:a novel source of heat tolerance at the vegetative and reproductive stages in rice[J]. Physiologia plantarum,2015,154(4):543-559.
(责任编辑:胡立萍)
