三种典型森林雉类的占域模式及影响因子
2025-01-12郭艳许兴如蒲冠桦尹华康付励强黄耀华白文科王彬张晋东周材权

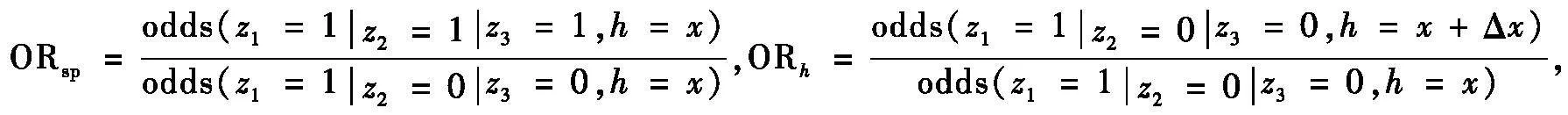
摘要:凉山山系马边大风顶国家级自然保护区山地森林内共存着多样性极为丰富的雉类物种,为了了解这些雉类的占域率、种间相互作用及其影响因子,于2021年5月至2023年6月,在该保护区内布设了131台红外相机,累计工作37 496相机日,对3种典型森林雉类红腹角雉(Tragopan temminckii,独立有效拍摄n=593)、血雉(Ithaginis cruentus,n=227)、白鹇(Lophura nycthemera,n=81)进行了监测,并使用多物种占域模型分析不同因子对其占域的影响。结果显示:3种雉类的占域率受不同环境变量的影响,即红腹角雉的占域率与海拔、坡度、草本盖度和灌木均高呈正相关,与距水源距离、人类活动强度和放牧强度呈负相关;血雉的占域率与坡向、坡度、海拔、距水源距离、乔木均高、灌木均高和草本盖度呈正相关,与人类活动强度呈负相关;白鹇的占域率与坡向呈正相关,与海拔、坡度、乔木均高、灌木均高、草本盖度和放牧强度呈负相关。红腹角雉和血雉之间具有负相互作用,红腹角雉和白鹇、血雉和白鹇之间具有正相互作用。3种雉类两两之间的相互作用对其占域率影响的重要性均大于栖息地特征和人为干扰。综上,种间相互作用、栖息地特征和人为干扰共同影响了凉山山系3种森林雉类的占域和共存。基于研究结果,建议保护区在开展多物种整合保护时,加强对物种之间相互作用的评估和对雉类栖息地内的人为活动和干扰的管控。
关键词:多物种占域模型;鸡形目;种间相互作用;人为干扰;四川马边大风顶国家级自然保护区
中图分类号:Q958.1文献标志码:A文章编号:1673-5072(2025)01-0001-11
Occupancy Pattern and Influencing Factorsof" Three Representative Forest Pheasants
Abstract:There are diverse pheasant species coexisting in the mountainous forests of Mabian Dafengding National Nature Reserve in Liangshan Mountains.In order to understand the occupancy rate,interspecific interactions and influencing factors of these pheasants,131 infrared-triggered cameras were set up there from May 2021 to June 2023,with a total of 37 496 camera-trapping days,to monitor the three representative forest pheasants,namely the Temminck’s tragopan (Tragopan temminckii,independent capture n=593),the blood pheasant (Ithaginis cruentus,n=227) and silver pheasant (Lophura nycthemera,n=81).The multi-species occupation model was applied to analyze the effects of different factors on the occupancy rate.The results show that the occupancy rate of the three pheasant species is influenced by different environmental variables.The results show that the occupancy rate of the Temminck’s tragopan is positively correlated with elevation,slope,herb coverage and mean shrub height,and negatively correlated with distance from water resources,human activity intensity and grazing intensity;the occupancy rate of the blood pheasant is positively correlated with aspect,slope,elevation,distance from water resources,mean tree height,mean shrub height and herb coverage,and is negatively correlated with human activity intensity;the occupancy rate of the silver pheasant is positively correlated with aspect,and negatively correlated with elevation,slope,mean tree height,mean shrub height,herb coverage and grazing intensity.There is a negative interaction between Temminck’s tragopan and blood pheasant,whereas a positive interaction between Temminck’s tragopan and silver pheasant,as well as between blood pheasant and silver pheasant.The influence of the pairwise interactions between three pheasants on their occupancy rate is greater than that of habitat characteristics and human interference.In summary,interspecific interactions,habitat characteristics and human disturbances jointly affect the occupancy and co-existence of forest pheasants in Liangshan Mountains.Based on our findings,it is suggested that the evaluation of species interaction and the control of human activities in pheasant habitats should be strengthened when multi-species integrated conservation is carried out in nature reserves.
Keywords:multi-species occupancy model;Galliformes;interspecific interaction;human disturbance;Sichuan Mabian Dafengding National Nature Reserve
占域率是指栖息地中某一个区域或斑块被目标物种占据和使用的概率[1],栖息地质量、资源丰富度[2]、种间相互作用[3-4]、捕食风险[5]、人为干扰[6]等生态因子以及各生态因子之间的交互作用[7]都可能会驱动物种的占域状态发生变化。物种之间的相互作用在动物群落的构建中发挥着重要作用,对于生态位相似的物种,种间竞争可能会降低其中至少一个物种的适合度[3],从而改变其占域模式。同时,日益加剧的人类活动也可能对物种的栖息地利用产生显著影响,野生动物倾向于通过改变自身的栖息地使用或活动模式来应对人为干扰,以在空间或时间上回避人类[8-9]。当群落中共存着多个具有相互作用的物种时,人为干扰的影响和作用机制则会更加复杂。人类活动对栖息地的干扰类型和强度在空间分布上并不均匀,造成了不同程度的栖息地破碎化[10]。一方面,这种破碎化会压缩物种已有的栖息地,将具有相似栖息地需求的物种限制到更狭窄的生态位空间之中,并因此产生更激烈的种间竞争[11-12],甚至是竞争排斥[13]。另一方面,不同物种对于干扰的耐受程度有差异[6],在相同干扰强度的影响下耐受程度不同的2个物种可能会选择不同的栖息地[14],从而减弱种间竞争并实现同域物种之间的共存。
雉科(Phasianidae)鸟类体型大、飞行和扩散能力弱,是典型的地栖鸟类。但由于孵化期和性成熟时间长、对环境变化和人为干扰敏感、躲避天敌能力弱等特点[15],导致与其他鸟类相比,雉类数量增长缓慢,更易濒危[16]。我国分布有28属64种雉类,其中51种被列为国家重点保护动物[17]。有研究表明,放牧[18]、砍伐森林[19]、旅游[20]、道路和土地开发[10]、偷猎[21]、采药和采笋[20]等人为干扰导致雉类栖息地急剧缩减和破碎化以及种群数量迅速减少,是野生雉类的重要威胁。然而,不同类型和强度的人为干扰如何影响同域分布雉类物种的栖息地选择和种间相互作用的研究相对较少。Li等[2]研究发现,褐马鸡(Crossoptilon mantchuricum)喜欢在远离人类活动的地方觅食,雉鸡(Phasianus colchicus)却总是选择离人类活动较近的地方;李伟[21]对同域分布的黑颈长尾雉(Syrmaticus humiae)和白鹇(Lophura nycthemera)的夜栖地利用比较研究发现,白鹇对人为干扰的适应能力比黑颈长尾雉相对较强;吕盛寒[14]研究发现,白鹇和黄腹角雉(Tragopan caboti)的栖息地分化体现在白鹇会出现在竹林茶园等边缘有人类活动的地方,而黄腹角雉则不在这几种生境类型内活动。因此,研究人为干扰对雉类栖息地选择的影响,不但有助于回答同域分布雉类的共存模式,而且有利于雉类物种数量的保护和多物种的整合保护。
凉山山系地处四川省西南部,横断山区东南,属于全球36个生物多样性热点之一的中国西南山地[22]。由于特殊的地理位置和气候条件,凉山山系保存了面积巨大的亚热带山地森林生态系统,其中共存着多样性极为丰富的雉类物种[23]。马边大风顶国家级自然保护区位于凉山山系中段,是凉山山系保护区网络的重要组成部分[23],保护区内有多种雉类物种,其中红腹角雉(Tragopan temminckii)、血雉(Ithaginis cruentus)和白鹇在保护区内种群密度大,是保护区内的优势森林雉类物种,且均为国家Ⅱ级重点保护野生动物,具有较强的代表意义[24]。这3种雉类亲缘关系较近,都主要是地面取食,均以植食性食物为主,辅以少量的动物性食物[25-27]。关于这3种雉类的栖息地利用已有一些研究[28-31],但关于它们在同域分布时的种间相互作用和不同环境变量对栖息地利用的影响还未见报道。因此,本研究在四川马边大风顶国家级自然保护区使用红外相机监测技术和多物种占域模型对红腹角雉、血雉、白鹇3种典型森林雉类的占域模式进行研究,以期探讨:(1)同域共存的3个雉类物种的占域率及其影响因子;(2)种间相互作用和环境协变量的相对重要性。
1材料和方法
1.1研究区域
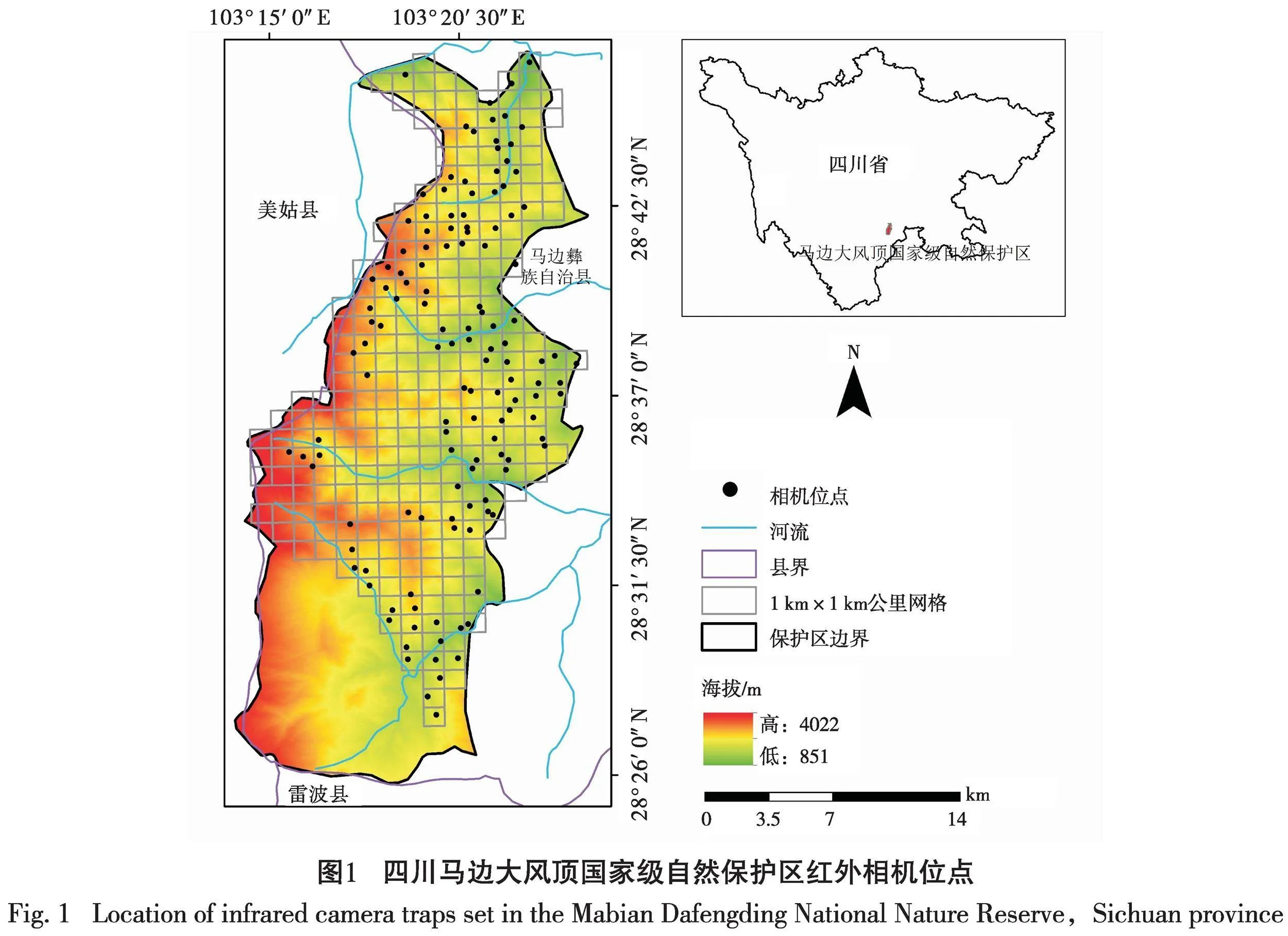
四川马边大风顶国家级自然保护区位于四川省乐山市马边彝族自治县,地势由西向东倾斜(图1),是目前世界上亚热带山地动植物资源及生境保存最完整的地区之一[24]。保护区内地表起伏大,气候垂直变化显著,自然植被分带现象明显,雨日多达240 d左右,年降水量1800~2000 mm[32]。保护区内具有丰富的动植物资源,如大熊猫(Ailuropoda melanoleuca)、羚牛(Budorcas taxicolor)、珙桐(Davidia involucrata)、水青树(Tetracentron sinense)等国家级珍稀濒危野生动植物,具有很高的研究价值与保护意义[33]。而且,保护区周边主要以彝族为主,具有极强的传统观念和生活习惯,对自然资源依赖较强,常通过放牧、采笋等人为活动来满足生活需求,对保护区内的动植物存在较大的影响[33-35]。
1.2红外相机布设
2021年5月至2023年6月,采用红外相机技术在四川马边大风顶国家级自然保护区展开监测。将调查区域划分为若干个1 km×1 km的网格(图1),根据野生动物的足迹、粪便和取食痕迹等选取单元网格,在单元网格中动物经常活动的走廊生境或靠近水源的地带放置1台相机,记录每台红外相机的GPS位点和海拔等信息,校对日期和时间,参数设置均为拍摄模式,连拍3张照片并录10 s的视频,灵敏度中等,采用24 h监测。本研究一共安放了131台相机。
1.3环境协变量
根据研究区域实际情况以及以往研究,选取可能对雉类栖息地利用产生影响的环境因子作为环境协变量。选择海拔(Ele)、坡向(Asp)、坡度(Slo)、乔木均高(Trh)、乔木平均胸径(Dbh)、乔木郁闭度(Trc)、灌木盖度(Shc)、灌木均高(Shh)、草本均高(Gsh)、草本盖度(Gsc)、距水源距离(Dtw)、放牧强度(Gtr)和人类活动强度(采笋、挖药、砍伐和巡护等人员)(Hmr)等13种为占域协变量;将每个探测周期中具体探测天数(Day)作为探测协变量。坡度和坡向均从地理空间数据云(http://www.gscloud.cn)下载的大风顶国家级自然保护区GDEMV3 30 m分辨率数字高程数据中获得。河流数据从全国地理信息资源目录服务系统(https://www.webmap.cn)中下载,并使用ArcGIS 10.7计算每个样点到最近河流的欧氏距离。基于每个相机位点拍摄到的牲畜(牛、羊等)和人类活动的独立照片数量计算相对多度(RAI),分别表征放牧强度和人类活动强度[36-37]。在模型构建过程中,将所有占域协变量数据标准化到0和1之间,以压缩变量离散程度。为避免协变量之间的多重共线性引起结果偏差,计算协变量的Pearson相关系数,若相关系数的绝对值小于0.5,则表明2个变量之间不存在较强的共线性,可以放入同一模型[38];若大于0.5,则删除其中1个协变量,选择保留生态学意义较大的协变量用于下一步分析。
1.4多物种占域模型
Rota等[39]的多物种占域模型是单季节占域模型[40]的推广,可以适应2个或多个相互作用的物种。多物种占域模型假设潜在的占用状态是一个多元伯努利随机变量,它允许构建数值稳定的影响物种共现的协变量模型,不需要不对称相互作用的先验假设[39];模型可以使用基本概率论在结果中获得物种两两之间的协方差或相关系数[39];该模型还允许将2个或多个物种占据同一地点的概率作为协变量函数进行建模[39]。在构建模型的过程中,本研究将每个调查地点的相机捕获历史以30 d划分为1个探测周期,共25个探测周期,当物种在1个探测周期中出现1次及以上记为1,否则记为0,数据缺失用NA代替补齐。使用两步法来降低多物种模型的复杂性[41]。
第一步,使用R 4.2.3[42]中的“unmarked”包[43-44]构建一系列的单物种占域模型,对所有协变量进行组合运算,采用MuMIn包[45]对所有候选模型的基于小样本量修正的赤池信息准则(Akaike Information Criterion Corrected for Small Sample Size,AICc)进行排序[46],并计算模型权重。将ΔAICc≤2的所有模型均视为等效模型,选取等效模型中的占域和探测协变量作为每个物种的最佳占域和探测协变量。
第二步,设计3组多物种占域模型,纳入上述过程筛选出的每个物种的最佳占域和探测协变量,每组模型设置不同的种间占域关系假设:(1)假设各物种的栖息地占域率独立于其他物种,只反映为占域协变量的函数(假设M1);(2)假设物种间具有恒定关系,即各物种的占域率不仅反映与占域协变量的函数关系,还受恒定不变的种间相互作用的影响(M2);(3)假设各物种的占域率与人为干扰因子的函数关系随着其他物种的出现与否变化,即种间相互作用与人为干扰因子间存在交互作用(M3)。使用“unmarked”包以惩罚似然函数构建多物种模型[47],计算模型的AIC(Akaike Information Criterion)值[46],根据3组假设构建的所有模型中最优模型(AIC值最小)的种间关系假设、重要协变量及其系数推断不同因子对物种栖息地选择和分化的影响。最优模型(AIC值最小)中每个协变量的回归系数(β)反映了目标物种的栖息地选择情况[1,39],β>0表示物种的占域率与该环境协变量呈正相关,β<0表示物种的占域率与该环境协变量呈负相关。
本研究对影响3种雉类占域的多个协变量计算了当协变量(h)保持在基线水平(x)时,其他物种存在与否和每个物种占域率的优势比(ORsp)和每个物种的占域率与协变量的每一单位变化的优势比(ORh)的绝对对数优势比(|log(ORsp)|和|log(ORh)|),以衡量环境变量和种间相互作用对每个物种占域率的相对重要性[7, 39]。公式为
式中:odds为优势比(Odds Ratio),即某个物种占据某个场地的概率除以其不占据该场地(或者其在协变量(h)发生一个单位变化时占据该场地)的概率;z为物种;x为协变量h的每一单位变化值。
2结果
2.1最优模型筛选
2021年5月—2023年6月,131个监测位点累计工作37 496相机日,分别记录到红腹角雉、血雉和白鹇的独立有效照片593张、227张和81张。通过单物种占域模型,红腹角雉、血雉和白鹇分别获得9个等效模型,识别出各物种最佳占域和探测协变量(表1):红腹角雉、血雉和白鹇的探测都受每个探测周期中具体探测天数的影响。红腹角雉的最佳占域协变量有海拔、坡度、灌木均高、草本盖度、距水源距离、人类活动强度和放牧强度7个;血雉的最佳占域协变量有海拔、坡向、坡度、乔木均高、灌木均高、草本盖度、距水源距离和人类活动强度8个;白鹇的最佳占域协变量有海拔、坡向、坡度、乔木均高、灌木均高、草本盖度和放牧强度7个。
将上述影响3种雉类占域率的协变量纳入多物种占域模型中,根据上述3个假设共拟合出5个模型(表2),其中假设M1(M1-1)和假设M2(M2-1)各构建1个模型,假设M3构建3个模型(M3-1、M3-2和M3-3)。模型M1-1反映了3种雉类独立出现,且每个物种占域率受占域协变量的影响;模型M2-1反映了3种雉类之间具有恒定不变的种间相互作用,且每个物种占域率受占域协变量和种间相互作用的影响;模型M3-1反映了人类活动强度和放牧强度与3种雉类的占域率之间的关系因其他物种的存在与否而异;模型M3-2反映了人类活动强度与3种雉类的占域率之间的关系因其他物种的存在与否而异;模型M3-3反映了放牧强度与3种雉类的占域率之间的关系因其他物种的存在与否而异。根据表2可知,AIC值最小的最优模型为M2-1。
2.2各协变量的影响
最优模型(M2-1)显示,每个物种的占域率受到栖息地特征和人为干扰的影响。红腹角雉的占域率随着海拔(β=3.380)、坡度(β=1.501)、草本盖度(β=0.748)和灌木均高(β=2.601)的增加而增加,随着距水源距离(β=-1.474)、人类活动强度(β=-9.239)和放牧强度(β=-3.631)的增加而减小;血雉的占域率随着坡向(β=0.285)、坡度(β=2.965)、海拔(β=16.053)、距水源距离(β=2.040)、乔木均高(β=2.033)、灌木均高(β=0.882)和草本盖度(β=1.069)增加而增加,随着人类活动强度(β=-4.385)的增加而减少;白鹇的占域率随坡向(β=0.951)的增加而增加,随着海拔(β=-18.051)、坡度(β=-5.233)、乔木均高(β=-5.687)、灌木均高(β=-0.656)、草本盖度(β=-3.218)和放牧强度(β=-4.542)的增加而减小(图2)。
2.3种间相互作用和协变量的相对重要性
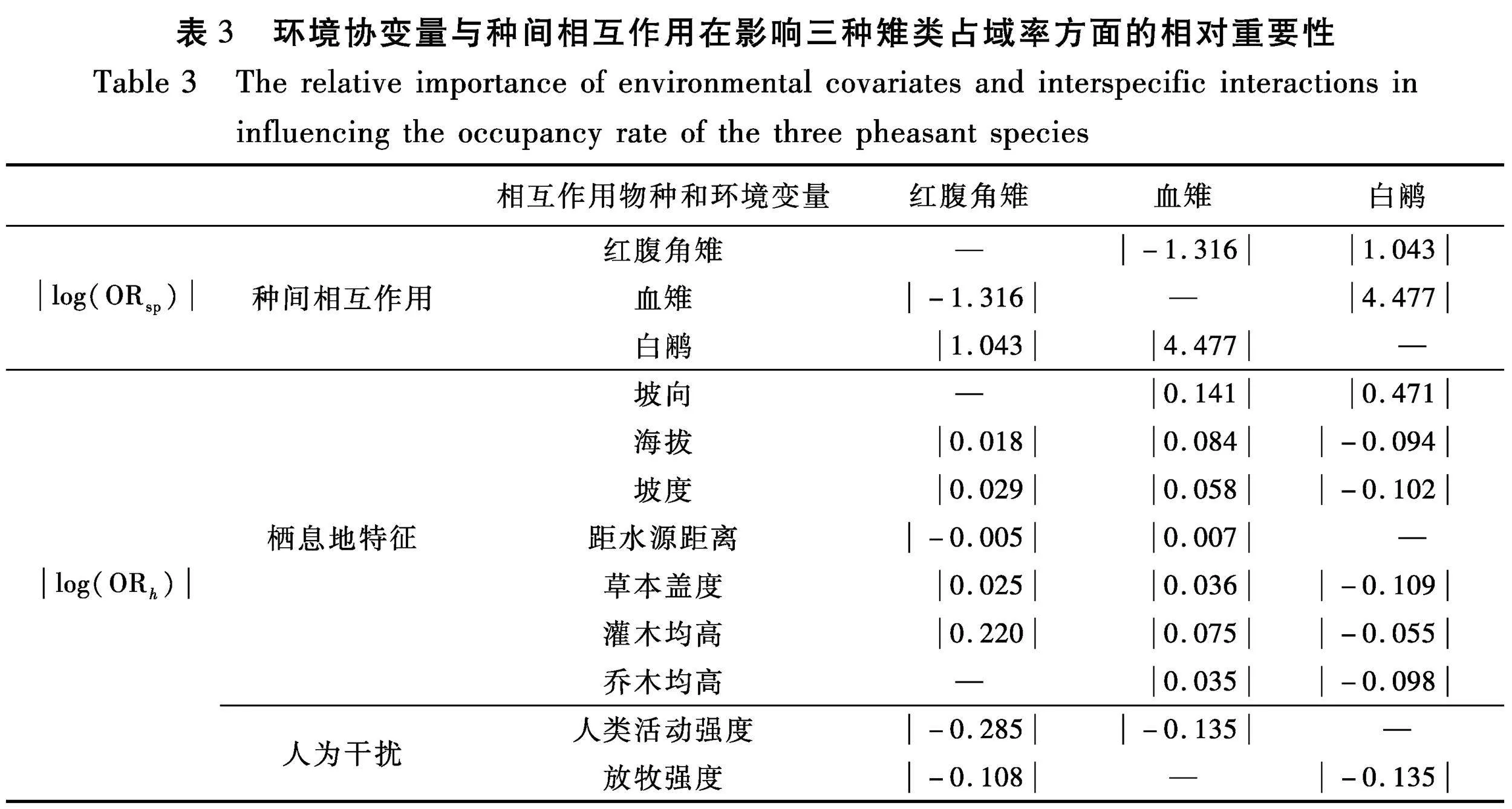
基于最优模型(M2-1)计算ORsp与ORh的绝对对数优势比,确定影响3种雉类占域率的不同占域协变量和种间相互作用的相对重要性。红腹角雉和血雉之间具有负相互作用,而红腹角雉和白鹇、血雉和白鹇之间具有正相互作用,3种雉类两两之间的相互作用对物种占域率影响的相对重要性均大于坡向、海拔、坡度、距水源距离、人类活动强度、放牧强度、草本盖度、灌木均高和乔木均高等9种环境因子(表3)。除种间相互作用外,在栖息地特征和人为干扰变量中灌木均高和人类活动强度是影响红腹角雉占域率的重要因子;坡向和人类活动强度是影响血雉占域率的重要因子;坡向和放牧强度是影响白鹇占域率的重要因子(表3)。
3讨论
本研究通过长期监测和综合分析,首次揭示了凉山山系森林生态系统中3种代表性雉类物种(红腹角雉、血雉、白鹇)的占域模式。乔木均高、灌木均高和草本盖度等不同的植被特征反映了栖息地内隐蔽条件和食物资源的差异,影响着不同雉类物种的栖息地选择和占域[48]。本研究显示,乔木均高、草本盖度和灌木高度等植被特征对3种雉类的占域影响具有较大差异,表明这3种雉类会倾向选择具有不同植被特征的栖息地来促进它们之间的共存。高大的乔木和灌木可以为雉类物种提供良好的隐蔽条件,增加雉类在林下层中取食的安全性,同时还提供了适宜的夜栖场所,但是过高的乔木和灌木会影响林下植被的生长状况,从而影响林下植被的资源丰富度[49]。权衡利弊,白鹇的占域率随着乔木均高和灌木均高的增大而减小;而红腹角雉由于取食种类广泛[27],且保护区良好的灌木条件已经满足了其躲避天敌和夜栖的条件,其占域率随着灌木均高的增大而增大,却不受乔木高度的影响;血雉主要生存在高海拔区域,温度较低,高大的乔木和灌木不但可以涵养水源,还可以抵御部分雨雪,有利于血雉保存热量和觅食[50],因此血雉的占域率随着乔木均高和灌木均高的增大而增大。3种雉类均以植物性食物为主,兼食动物性食物,草本除了本身的茎叶可供雉类取食外,还为多种昆虫的生存提供了良好的条件[18],即草本盖度越大食物资源可能更丰富,因此红腹角雉和血雉的占域率随着草本盖度的增大而增大,而白鹇雄鸟具较长的尾羽,过于茂密的草本会导致其穿行困难,阻碍躲避危险的速度[30],权衡避敌和取食,白鹇的占域率随着草本盖度的增大而减小。
血雉和白鹇的占域率均随着坡向的增大而增大,这可能是由于阴坡土壤较湿润,肥力较高,腐殖质含量较多,具有较高的植物多样性,能为雉类提供较好的食物资源[51];而红腹角雉的占域率不受坡向的影响,这可能是由于红腹角雉的食性较广泛,不同的坡向都能满足其取食的需求[29]。雉类对坡度的选择与体型大小、活动能力和取食有关[48,52]。红腹角雉和血雉的占域率随着坡度的增大而增大,可能是由于较大的坡度能够提供更多的斜坡,增大其进行滑翔逃亡的可能[30];而白鹇的占域率随着坡度的增大而减小,可能是由于白鹇体型较大,较长的尾羽还增加了额外的重量,它的敏捷性相较于体型较小的红腹角雉和血雉更弱,逃避天敌时需要更多的时间和空间[53],而且当遇到惊吓时白鹇并不立即起飞而是急步奔逃[54],因此白鹇会偏好于选择较为平缓的坡度。红腹角雉和血雉的占域率随着海拔的上升而增加,白鹇的占域率随着海拔的上升而减小。白鹇和血雉分别在低海拔带和高海拔带有最大的占域率,而红腹角雉作为优势物种,在中到高海拔带均有较高的占域率,这种垂直水平生态位上的分离,缓和了3种雉类在空间生态位上的竞争。
红腹角雉和血雉之间有负相互作用,即潜在的种间竞争关系[55],二者在栖息地选择上存在较大的重叠,同时也在某些方面表现出分离,这种分离暗示了栖息地分化可能是红腹角雉和血雉减弱种间竞争,实现同域共存的重要方式。如,红腹角雉和血雉的占域率在海拔上存在很大的差异,它们在海拔上的垂直分布特征体现出了两种雉类在海拔生态位上的分离,减少了两种雉类在食物资源和其他资源上的竞争,促进了两种雉类长时间的同域共存[29]。红腹角雉和白鹇、血雉和白鹇之间有正相互作用,在相同的栖息地条件下,种间正相互作用可能对至少一个物种产生积极影响[55]。红腹角雉和白鹇、血雉和白鹇两两之间在栖息地选择上表现出较大的分离,偏好不同的栖息地特征,增加了他们同域共存的适合度。
人为干扰对野生动物的影响取决于干扰的类型和强度,并且通常因物种而异[8-9]。家畜的觅食和踩踏可能会减少植被的异质性,降低节肢动物的多样性和丰富度[56],因此,放牧强度的增大通常暗示着食物资源的减少,红腹角雉和白鹇的占域率受放牧强度的消极影响,且放牧强度是栖息地特征和人为干扰因子中影响白鹇占域的第二重要因子。本研究区域的放牧主要集中在中低海拔地区,而血雉的占域率随海拔的升高而增大,因此血雉的占域受放牧的影响较小。人类活动可能会导致鸟类的适宜栖息地大面积减少或种群数量急剧下降等[57]。红腹角雉和血雉的占域均回避人类活动,且人类活动强度还是栖息地特征和人为干扰因子中影响红腹角雉的最重要因子和影响血雉占域的第二重要因子,表明人类活动对于两者而言是一种较强的干扰和威胁。相反,白鹇的占域率不受人类活动强度的影响,可能是由于占域模型的构建考虑的是所有协变量对白鹇占域的综合影响,而白鹇对人为干扰具有较强的适应能力[14,21],其他协变量对白鹇占域的较大影响可能掩盖了人类活动强度对其较小的影响。
综上所述,凉山山系山地森林雉类的占域和共存模式不但受到栖息地特征和人为干扰的影响,还与群落中物种之间的相互作用有关。本研究发现3种雉类倾向于选择具有不同特征的栖息地,因此,它们的共存对栖息地异质性具有较高的要求。当进一步考虑同域分布的四川山鹧鸪(Arborophila rufipectus)、白腹锦鸡(Chrysolophus amherstiae)等其他雉类物种的栖息地需求时,栖息地异质性的重要性可能会更加明显,所以保护区在未来的保护工作中应加强对景观和微生境尺度的栖息地异质性的维持。放牧和人类活动是保护区内最普遍的两种干扰类型,它们对森林雉类栖息地占域的影响都较为重要,并且主要体现为消极影响。因此,建议保护区进一步加强对放牧的管控,减轻家畜对植被和同域动物的影响。此外,根据红外相机监测结果,除了进山放牧的人外,还有进山采药和采笋的人,因此,在未来的保护工作中,需严格控制居民上山采摘,并为当地社区创造更多的环境友好型就业机会,以减缓保护物种和人类生产生活之间的矛盾,降低人类干扰对野生动物栖息地的破坏。
致谢:感谢西弗吉尼亚大学(West Virginia University)和密苏里大学哥伦比亚分校(University of Missouri Columbia)的Christopher Rota、北卡罗来纳自然科学博物馆(North Carolina Museum of Natural Sciences)和北卡罗莱纳州立大学(North Carolina State University)的Arielle Parsons对多物种占域模型使用的指导,感谢四川马边大风顶国家级自然保护区保护中心和西华师范大学所有参与野外工作人员的辛苦付出。
参考文献:
[1]肖文宏,束祖飞,陈立军,等.占域模型的原理及在野生动物红外相机研究中的应用案例[J].生物多样性,2019,27(3):249-256.
[2]LI H Q,LIAN Z M,CHEN C G.Winter foraging habitat selection of brown-eared pheasant (Crossoptilon mantchuricum) and the common pheasant (Phasianus colchicus) in Huanglong Mountains,Shaanxi Province[J].Acta Ecologica Sinica,2009,29(6):335-340.
[3]WANG F,MCSHEA W J,WANG D J,et al.Shared resources between giant panda and sympatric wild and domestic mammals[J].Biological Conservation,2015,186:319-325.
[4]CT I M,DARLING E S,BROWN C J.Interactions among ecosystem stressors and their importance in conservation[J].Proceedings of the Royal Society B:Biological Sciences,2016,283(1824):20152592.
[5]HOPCRAFT J G C,ANDERSON T M,PREZ-VILA S,et al.Body size and the division of niche space:food and predation differentially shape the distribution of Serengeti grazers[J].Journal of Animal Ecology,2012,81(1):201-213.
[6]夏珊珊,胡大明,邓玥,等.同域分布红腹锦鸡和红腹角雉在不同空间尺度下的生境分化[J].生态学报,2019,39(5):1627-1638.
[7]PARSONS A W,ROTA C T,FORRESTER T,et al.Urbanization focuses carnivore activity in remaining natural habitats,increasing species interactions[J].Journal of Applied Ecology,2019,56(8):1894-1904.
[8]GAYNOR K M,HOJNOWSKI C E,CARTER N H,et al.The influence of human disturbance on wildlife nocturnality[J].Science,2018,360(6394):1232-1235.
[9]LAMRIS D W,TAGG N,KUENBOU J K,et al.Drivers affecting mammal community structure and functional diversity under varied conservation efforts in a tropical rainforest in Cameroon[J].Animal Conservation,2020,23(2):182-191.
[10]石江艳.人为干扰下白冠长尾雉及其同域分布的兽类活动节律分析[D].北京:北京林业大学,2020.
[11]HAGEN C A,PITMAN J C,ROBEL R J,et al.Niche partitioning by lesser prairie-chicken Tympanuchus pallidicinctus and ring-necked pheasant Phasianus colchicus in southwestern Kansas[J].Wildlife Biology,2007,13:34-41.
[12]邓文洪.栖息地破碎化与鸟类生存[J].生态学报,2009,29(6):3181-3187.
[13]ROBINSON S K,TERBORGH J.Interspecific aggression and habitat selection by Amazonian birds[J].Journal of Animal Ecology,1995,64(1):1-11.
[14]吕盛寒.基于红外相机技术对福建武夷山国家自然保护区白鹇(Lophura nycthemera)、黄腹角雉(Tragopan caboti)时空利用行为的研究[D].厦门:厦门大学,2019.
[15]张正旺,丁长青,丁平,等.中国鸡形目鸟类的现状与保护对策[J].生物多样性,2003,11(5):414-421.
[16]郑光美,王岐山.中国濒危动物红皮书 鸟类[M].北京:科学出版社,1998.
[17]郑光美.中国鸟类分类与分布名录[M].4版.北京:科学出版社,2023.
[18]李伟,周伟,刘钊,等.云南大中山黑颈长尾雉栖息地选择周年变化[J].动物学研究,2010,31(5):499-508.
[19]诸葛阳,丁平.浙江省珍稀雉类的分布生境和资源保护[J].野生动物学报,1988,9(4):3-4.
[20]钟雪,邓玥,杨彪,等.四川白水河国家级自然保护区红腹锦鸡种群密度及微生境选择[J].四川大学学报(自然科学版),2020,57(1):174-180.
[21]李伟.云南大中山黑颈长尾雉栖息地选择与种间栖息地利用比较[D].昆明:西南林学院,2007.
[22]MITTERMEIER R A,GIL P R,HOFFMANN M,et al.Hotspots revisited.Earth’s biologically richest and most endangered terrestrial ecoregions[M].Mexico:Cemex,2004.
[23]黄钟宣.凉山山系地区野生植物调查与景观利用评价[D].成都:成都理工大学,2021.
[24]吴沛桦,侯金,黄耀华,等.利用红外相机调查马边大风顶保护区鸟类资源的时空分布特征[J].四川林业科技,2020,41(1):27-35.
[25]史东仇,李贵辉.四川南坪白河保护区血雉食性的初步研究[J].动物学研究,1985,6(2):139-145.
[26]高育仁.白鹇食性研究[J].动物学研究,1993,14(2):194-196.
[27]史海涛,郑光美.红腹角雉的食性研究[J].动物学研究,1998,19(3):225-229.
[28]李伟,周伟,张兴勇,等.哀牢山国家级自然保护区南华片三种雉类春季取食地利用比较[J].动物学研究,2006,27(5):495-504.
[29]崔鹏,康明江,邓文洪.繁殖季节同域分布的红腹角雉和血雉的觅食生境选择[J].生物多样性,2008,16(2):143-149.
[30]李仁贵,黄金燕,周世强,等.卧龙大熊猫栖息地红腹角雉冬季生境选择的研究[J].四川林业科技,2011,32(2):55-59.
[31]FAN F,BU H L,MCSHEA W J,et al.Seasonal habitat use and activity patterns of blood pheasant Ithaginis cruentus be in the presence of free-ranging livestock[J].Global Ecology and Conservation,2020,23:e01155.
[32]毛泽恩,白文科,付励强,等.马边大风顶自然保护区分布存疑兽类调查[J].西华师范大学学报(自然科学版),2022,43(1):1-8.
[33]吴灵芝,曲别曲日,蒋平,等.四川马边大风顶自然保护区周边社区对当地自然保护的影响[J].四川动物,2007,26(4):881-883.
[34]尹华康,张晋东,黄金燕,等.四川马边大风顶自然保护区大熊猫主食竹空间分布特征[J/OL].南京林业大学学报(自然科学版),1-9[2024-02-03].http://kns.cnki.net/kcms/detail/32.1161.s.20230726.1053.002.html.
[35]舒习玉,黄耀华,贺俊东,等.不同禁牧措施对高山草甸植物群落的影响[J].西华师范大学学报(自然科学版),2021,42(2):105-109.
[36]LI S,MCSHEA W J,WANG D J,et al.The use of infrared-triggered cameras for surveying phasianids in Sichuan Province,China[J].IBIS,2010,152(2):299-309.
[37]GU B J,WENG Y,DIAO Y X,et al.Is livestock grazing compatible with green peafowl (Pavo muticus) conservation?Potential chance of peafowl-human coexistence[J].Biological Conservation,2022,275:109772.
[38]TAN C K W,ROCHA D G,CLEMENTS G R,et al.Habitat use and predicted range for the mainland clouded leopard Neofelis nebulosa in Peninsular Malaysia[J].Biological Conservation,2017,206:65-74.
[39]ROTA C T,FERREIRA M A R,KAYS R W,et al.A multispecies occupancy model for two or more interacting species[J].Methods in Ecology and Evolution,2016,7(10):1164-1173.
[40]MACKENZIE D I,NICHOLS J D,LACHMAN G B,et al.Estimating site occupancy rates when detection probabilities are less than one[J].Ecology,2002,83(8):2248-2255.
[41]ANDRADE-PONCE G P,MANDUJANO S,DáTTILO W,et al.A framework to interpret co-occurrence patterns from camera trap data:the case of the gray fox,the bobcat,and the eastern cottontail rabbit in a tropical dry habitat[J].Journal of Zoology,2022,318(2):91-103.
[42]R CORE TEAM.R:a language and environment for statistical computing[CP/DK].R Foundation for Statistical Computing,Vienna,Austria,2023.https://www.R-project.org/.
[43]FISKE I,CHANDLER R.Unmarked:an R package for fitting hierarchical models of wildlife occurrence and abundance[J].Journal of Statistical Software,2011,43(10):1-23.
[44]KELLNER K F,SMITH A D,ROYLE J A,et al.The unmarked R package:twelve years of advances in occurrence and abundance modelling in ecology[J].Methods in Ecology and Evolution,2023,14(6):1408-1415.
[45]BARTON K.MuMIn:Multi-Model inference[CP/DK].R package version 1.47.5.2023.https://CRAN.R-project.org/package=MuMIn.
[46]AKAIKE H.A new look at the statistical model identification[J].IEEE Transactions on Automatic Control,1974,19(6):716-723.
[47]CLIPP H L,EVANS A L,KESSINGER B E,et al.A penalized likelihood for multispecies occupancy models improves predictions of species interactions[J].Ecology,2021,102(12):e03520.
[48]刘鹏.官山自然保护区四种雉类的生境选择[D].南京:南京林业大学,2011.
[49]王彬,朱博伟,徐雨,等.四川雉鹑(Tetraophasis szechenyii)的微生境选择及其季节性动态[J].生态学杂志,2022,41(6):1143-1148.
[50]徐雨,王彬,窦亮,等.三种同域高山鸡形目鸟类的生境选择[J].四川动物,2017,36(3):258-265.
[51]刘旻霞,南笑宁,张国娟,等.高寒草甸不同坡向植物群落物种多样性与功能多样性的关系[J].生态学报,2021,41(13):5398-5407.
[52]康明江,郑光美.白腹锦鸡雌鸟繁殖期的活动区和栖息地选择[J].北京师范大学学报(自然科学版),2007,43(5):558-562.
[53]方小斌,邹瑀琦,丁长青.鸟类惊飞距离及其影响因素[J].动物学杂志,2017,52(5):897-910.
[54]李炳华,陈壁辉.皖南白鹇的地理分布及生态初步调查[J].动物学杂志,1984,19(4):15-18.
[55]ZHANG T X,XILI Y Z,RAN J H,et al.Effects of environmental changes on interspecific interactions of three sympatric pheasants:a study based on long-term monitoring data[J].Ecological Indicators,2022,135:108532.
[56]EVANS D M,VILLAR N,LITTLEWOOD N A,et al.The cascading impacts of livestock grazing in upland ecosystems:a 10-year experiment[J].Ecosphere,2015,6(3):1-15.
[57]杨兰荣,洪明生,韦伟,等.四川省典型湿地秋冬季的水鸟多样性及其干扰因素[J].西华师范大学学报(自然科学版),2019,40(1):22-31.
