间作模式对油橄榄土壤养分及土壤微生物的影响
2024-12-27黄楚亦胡青赵敏陆斌张光华王猛




关键词:油橄榄;间作;土壤养分;微生物多样性;群落结构
油橄榄Oleaeuropaea,为木樨科Oleaceae木樨榄属Olea常绿小乔木,是世界四大木本油料植物之一。油橄榄原产于地中海地区,在我国的主要适生区包括金沙江干热河谷区、白龙江低山河谷区、长江三峡低山河谷区[1]。这些区域光热资源充沛,为油橄榄的生长提供了良好的自然条件。然而,这些区域多位于水土流失和养分流失严重的半山干旱区[2],土壤质量较差,地表植被稀疏,生态环境脆弱。此外,油橄榄幼林期长,种植4~5a后才开始开花结果,7a后才能达到盛产期,这在一定程度上限制了油橄榄产业的生产效率和生态效益。因此,改善油橄榄园土壤管理方式是当前油橄榄产业发展亟待解决的问题之一。间作是一种高效的经济林经营措施,是目前在国内外广泛应用的一种现代化可持续发展的土壤管理方式[3]。将不同生态位作物复合种植,不仅能充分利用林地空间资源,提高土地资源的利用效率,还能发挥间作作物的生态功能,改善土壤环境,对提高油橄榄园的土地利用率及其生态和经济效益具有深远的意义。
土壤微生物是土壤生态系统的关键组成部分,是土壤生态系统中较为活跃的组分之一,也是养分转化和物质循环等土壤生态过程中必不可少的生物因素[4],对维持土壤生态系统的稳定和健康起到重要作用。土壤养分是植物正常生长发育的基础[5],土壤有机质和养分主要来源于土壤微生物对凋落物以及动植物残体的分解,与土壤微生物多样性和丰度密切相关。间作可以通过改变植物生理特征及根系分泌特性间接影响土壤微生物多样性和群落结构,从而改善土壤微环境[6-7]。赵春梅等[8]发现,在橡胶林下间作王草、益智和咖啡作物有利于提高土壤微生物群落功能多样性,改善土壤生态环境。不同的间作作物的生理特征和生物学特性不同,其对土壤养分及土壤微生物的影响也是不同的,如杨宏伟等[9]发现在油橄榄林间作百喜草增加了土壤有机质含量,并能够有效改善根际土壤微环境,而严毅等[10]发现在油橄榄林下种植百合后林地土壤养分含量明显降低。因此,研究不同间作模式下油橄榄土壤养分变化及微生物动态,对选择适宜的油橄榄间作模式和促进我国油橄榄产业高效发展具有重要意义。
目前关于油橄榄间作经营方式的探索已取得一些成果,如油橄榄间作毛苕子、百喜草、红三叶等绿肥作物[11-13],以及在油橄榄林下种植大球盖菇、榆黄蘑、灵芝和小麦等作物[14-15]。这些研究内容集中于根系发育形态、光合特性、抗旱生理、经济产量等方面。然而,关于不同间作模式下油橄榄林地土壤养分变化、微生物多样性及其群落结构变化等的深入分析报道较为鲜见。本研究中以油橄榄单作为对照,以3种油橄榄间作模式为研究对象,分析了间作模式对油橄榄土壤养分含量、土壤微生物多样性和群落结构的影响,旨在为探索油橄榄高效栽培模式和改善油橄榄林地土壤生态环境提供参考。
1材料与方法
1.1试验区概况
研究地位于云南省丽江市永胜县六德乡玉水村油橄榄栽培示范园(100°86′E,26°57′N)。该园地处云南省北部金沙江中游亚热带干热河谷地带,平均海拔1991.7m,年平均气温16.2℃,年平均日照2403h,年降水量800.40mm。干、热是该地区典型的气候特征,降水主要集中在夏秋季节,冬春干旱,全年光照充足。示范园土壤以砂壤土为主,土层较厚。整个园区油橄榄种植面积超46.67hm2,在试验基地内选择坡向、海拔且水肥管理措施基本一致的油橄榄‘佛奥’品种种植区域进行试验,株行距为5m×7m。
1.2试验设计
油橄榄为2017年初种植的品种‘佛奥’。试验设4个处理,以油橄榄单作为对照(CK),其余为间作模式,分别为油橄榄林下间作滇黄精(FD)、紫花苜蓿草(FZ)、3种混播草(F3)。2018年,将紫花苜蓿草与3种混播草种(紫花苜蓿草、黑麦草、金鸡菊)撒播于油橄榄林地内,将滇黄精种植于油橄榄行间(株行距0.5m×1.0m)。试验期间油橄榄树的施肥、灌溉等管护措施保持一致,均按照常规方法统一进行管理。
1.3试验方法
1.3.1土壤样品采集
2023年5月,采用典型的“S”形布设土壤取样法,在每个处理区域内设置3个土壤采样点。在距离油橄榄树干基部50cm处,除去地面的枯枝落叶、碎石和杂物,垂直采集深度0~10cm的土壤。每个样地采取3份土样,每处理重复3次,将其混合均匀为1份土样。将每份土壤样本取0.5kg,立即装入无菌自封袋并编号,置于冷冻箱中保存备用,用于土壤微生物群落结构及多样性的测定。将一部分土壤样品自然风干,用于养分含量等指标的测定。
1.3.2土壤理化指标测定[16]
土壤pH值采用电位计法(土、水体积比1∶2.5)测定,有机碳含量采用重铬酸钾容量法测定,全氮含量采用凯氏定氮法测定,速效钾和全钾含量采用火焰原子分光光度法测定,有效磷和全磷含量采用钼锑抗比色法测定,碱解氮含量采用碱解扩散法测定。
1.3.3土壤DNA的提取及微生物高通量测序
利用IlluminaMiseq高通量测序技术对细菌和真菌多样性进行分析。采用OMEGASoilDNAKit试剂盒提取土壤基因组DNA,利用1%琼脂糖凝胶电泳检测DNA质量,利用NanodropNC2000测定DNA浓度,具体操作步骤以自带说明书为标准。以提取的基因组DNA为模板,以细菌通用引物338F(5′-ACTCCTACGGGAGGCAGCA-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)对16S的V3~V4区进行PCR扩增,对土壤真菌ITS的V1区进行扩增,扩增引物为ITS5F(5′-GGAAGTAAAAGTCGTAACAAGG-3′)和ITS1R(5′-GCTGCGTTCTTCATCGATGC-3′)。利用IlluminaMiseq技术平台进行文库构建和Paired-end测序。
1.4数据处理与分析
使用Vsearch和Cutadapt软件进行去引物、拼接、质量过滤、去重、去嵌合体、聚类等流程,获取高质量序列,在97%相似度水平对高质量序列聚类,生成ASVs特征序列,并分别输出代表序列和OTU表格,最后去除OTU表格中丰度为1的OTU及其代表序列。使用Silva数据库和UNITE数据库比对ASVs特征序列,得到每个ASV对应的分类学信息。使用R脚本的VennDiagram包制作韦恩图,利用QIIME2分析平台对物种分类组成情况和物种多样性进行分析。采用SPSS22.0软件分析数据差异的显著性,采用Excel软件制作表格。
2结果与分析
2.1不同间作模式下土壤养分含量和pH值
由表1可知,4个处理下土壤的pH值为8.12~8.36,均为碱性土壤,FD、F3间作处理下土壤pH值相对较低,但处理间差异均不显著。相较于CK,FD间作模式下土壤有机碳、全氮、全磷、全钾的含量均有所降低,但并无显著差异,速效钾、有效磷、碱解氮含量分别显著降低了55.45%、45.55%和33.46%。相较于CK,FZ间作模式下土壤全氮、全磷含量无显著差异,间作后有机碳、速效钾和碱解氮含量分别显著降低了60.73%、56.01%和77.48%,全钾、有效磷含量显著增加了29.40%、49.87%,且含量均高于其他3个处理。相较于CK,F3间作模式下土壤全钾含量显著增加了24.59%,碱解氮含量有所降低但不显著,其他养分含量相较于CK均无显著差异,但含量均有所增加。综上可见,与对照组CK相比,3种油橄榄间作模式均能在一定程度上改变油橄榄林地土壤养分含量,FD和FZ间作处理下土壤部分速效养分含量显著降低,F3间作处理对提高油橄榄林地土壤养分含量的效果最佳。
2.2不同间作模式下土壤微生物群落状况
2.2.1微生物OTUs数量
由不同间作模式下油橄榄土壤微生物OTUs数量韦恩图可知(图1),4种间作模式下土壤细菌OTUs总数量为38652,真菌OTUs总数量为3059,共有细菌OTUs数量为1088,占OTUs总数量的2.81%,共有真菌OTUs数量为30,占OTUs总数量的0.98%。CK、FD、FZ、F3间作模式下土壤特有的细菌OTUs数量分别为5998、5304、4601、7851,分别占细菌特有OTUs总数量的25.25%、22.33%、19.37%、33.05%。CK、FD、FZ、F3间作模式下土壤特有的真菌OTUs数量分别为541、640、370、778,分别占真菌特有OTUs总数量的23.23%、27.48%、15.89%、33.40%。4种间作模式下土壤细菌和真菌所包含的OTUs数量和特有的OTUs数量均以F3间作模式最多,FZ间作模式最少。根据细菌的OTUs总数量和特有OTUs数量由大到小排列,各处理依次为F3、CK、FD、FZ;根据真菌的OTUs数量由大到小排列,各处理依次为F3、FD、CK、FZ。综上,与油橄榄单作CK相比较,不同处理间细菌和真菌OTUs数量均有一定的差别。F3处理模式能在一定程度上增加土壤中细菌、真菌OTUs数量,FZ处理降低了土壤细菌、真菌微生物OTUs数量。
2.2.2微生物α多样性指数
α多样性指数是反映微生物群落丰富度、多样性和均匀度的综合指标。从表2中多个多样性指数来看,4种间作模式下细菌和真菌多样性指数间无显著差异,细菌的Chao1、Shannon指数均大于真菌,各处理细菌和真菌多样性指数变化与OTUs数量变化趋势基本一致。根据细菌群落丰富度和多样性指数由大到小排列,各处理依次为F3、CK、FD、FZ;根据真菌群落丰富度和多样性指数由大到小排列,各处理依次为F3、FD、CK、FZ。
2.2.3微生物群落组成
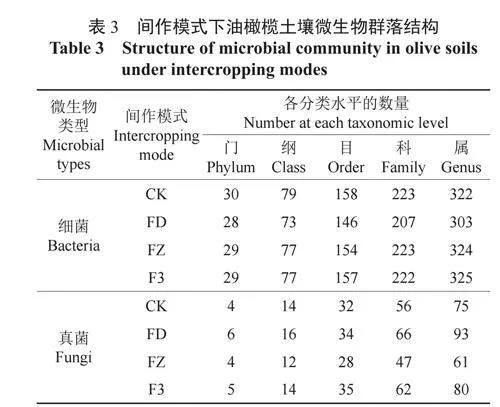
4种间作模式下油橄榄土壤细菌群落中共检测到33门83纲165目232科348属,真菌群落中共检测出7门17纲42目93科154属。由表3可知,细菌群落组成的数量在各分类水平上均明显高于真菌群落组成数量。根据细菌群落组成在各分类水平上由多到少排列,各处理依次为CK、F3、FZ、FD;根据真菌群落组成在各分类水平上由多到少排列,各处理依次为FD、F3、CK、FZ。
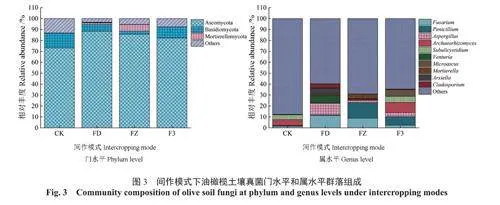
图2~3分别展示了4种间作模式下油橄榄土壤细菌和真菌在门和属分类学水平上相对丰度的变化规律,1种色块代表1个物种,色块的长度表示该物种的相对丰度。由图2A可知,4种间作模式下油橄榄土壤细菌门类相对丰度在前10位的菌群基本相同,分别为放线菌门Actinobacteriota、变形菌门Proteobacteria、酸杆菌门Acidobacteriota、绿弯菌门Chloroflexi、芽单胞菌门Gemmatimonadota、拟杆菌门Bacteroidota、黏球菌门Myxococcota、疣微菌门Verrucomicrobiota、Methylomirabilota、浮游球菌门Planctomycetota。4种间作模式下油橄榄土壤中优势菌门均为放线菌门(17.09%~31.03%)、变形菌门(21.27%~28.46%)、酸杆菌门(14.11%~16.60%)、绿弯菌门(8.95%~10.38%)、芽单胞菌门(7.06%~11.01%)。4种间作模式下放线菌门相对丰度由高到低分别为CK(31.03%)、FZ(29.72%)、F3(24.02%)、FD(17.09%),变形菌门相对丰度由高到低分别为FD(28.46%)、FZ(22.08%)、CK(21.84%)、F3(21.27%),酸杆菌门相对丰度由高到低分别为FD(16.60%)、CK(16.13%)、F3(15.58%)、FZ(14.11%)。4种间作模式下油橄榄土壤中细菌在属分类水平上的相对丰度具有一定差异(图2B),前3位优势菌属分别为Vicinamibacteraceae(4.03%~6.80%)、鞘氨醇单胞菌属Sphingomonas(2.97%~5.64%)、RB41(2.75%~4.67%),4种间作模式下Vicinamibacteraceae相对丰度由高到低分别为FD(6.80%)、CK(6.30%)、FZ(5.45%)、F3(4.03%),鞘氨醇单胞菌属相对丰度由高到低分别为FD(5.64%)、FZ(4.40%)、F3(3.20%)、CK(2.97%),RB41相对丰度由高到低分别为F3(4.67%)、FD(3.63%)、CK(3.16%)、FZ(2.75%)。综上,间作后各模式下土壤细菌在门、属水平群落物种组成上与CK相似,但间作后各菌群的相对丰度存在差异。
由图3A可知,间作模式下土壤优势真菌门类群主要为子囊菌门Ascomycota(88.42%~73.18%)、担子菌门Basidiomycota(2.87%~13.45%)。同时发现:被孢霉门Mortierellomycota为FZ间作模式下的优势菌门,相对丰度达6.03%,在其他模式中丰度较小。各间作模式按照土壤中子囊菌门的相对丰度由大到小排序,依次为FD(88.42%)、FZ(85.66%)、F3(82.20%)、CK(73.18%);按照担子菌门的相对丰度由大到小排序,依次为CK(13.45%)、F3(10.07%)、FD(6.21%)、FZ(2.87%)。由图3B可知,真菌属水平类群主要为镰刀菌属Fusarium、青霉属Penicillium、曲霉属Aspergillus、古根菌属Archaeorhizomyces、尖囊菌属Subulicystidium、黑星菌属Venturia、小囊菌属Microascus、被孢霉属Mortierella、阿泽拉霉属Arxiella、枝孢属Cladosporium。各间作模式下的优势菌属不同,CK模式下古根菌属(5.46%)和尖囊菌属(4.29%)是优势菌属,FD间作模式下镰刀菌属(11.32%)、曲霉属(10.52%)和黑星菌属(6.97%)为优势菌属,FZ间作模式下优势菌属为镰刀菌属(8.86%)和青霉属(14.36%),F3间作模式下土壤中优势菌属为古根菌属(9.26%)、青霉属(7.83%)、小囊菌属(6.13%)和尖囊菌属(5.66%)。综上,4种模式下土壤真菌在门水平上子囊菌门均为绝对优势门,且间作后子囊菌门的相对丰度较CK均有所增加,在属水平上各模式间优势属组成及相对丰度差异较大,3种间作模式均使部分土壤功能真菌微生物的相对丰度明显增加,其中FD和FZ模式增加了常见致病菌属镰刀菌属的相对丰度,病原菌属黑星菌属、枝孢属在FD模式下的相对丰度也较高,说明该间作模式可能会增加土壤病害侵扰的概率。
2.3不同间作模式下土壤微生物群落组成与土壤环境因子的相关性
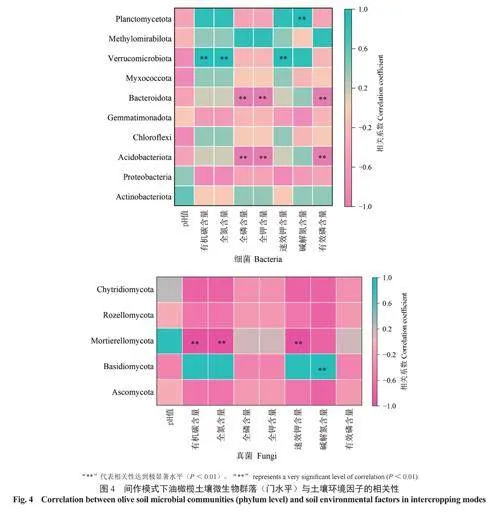
为了进一步了解土壤环境因子与土壤细菌、真菌群落的关系,将土壤中主要优势细菌和真菌菌群的相对丰度与土壤环境因子进行相关性分析。由土壤细菌相对丰度与土壤环境因子的Spearman相关性热图(图4)可知,在门分类水平上,酸杆菌门和拟杆菌门的相对丰度与全磷含量、全钾含量、有效磷含量极显著负相关,疣微菌门的相对丰度与有机碳含量、全氮含量、速效钾含量极显著正相关,浮游球菌门的相对丰度与碱解氮含量极显著正相关。由土壤真菌相对丰度与土壤环境因子的Spearman相关性热图(图4)可知,担子菌门Basidiomycota的相对丰度与碱解氮含量极显著正相关,被孢霉门的相对丰度与有机碳含量、全氮含量、速效钾含量极显著负相关。同时,土壤pH值与细菌、真菌各类菌群的相对丰度均无显著相关性。综上,细菌和真菌群落结构与土壤环境因子有着密切的相关性,但影响其土壤微生物群落结构的主要环境因子存在差异。
3讨论与结论
3.1间作模式对土壤养分元素含量的影响
大量研究结果证明,间作会对土壤环境因子产生影响[17-19]。本研究中,相较于CK,3种间作模式均对土壤养分含量产生了影响,FD间作模式下土壤速效钾、有效磷、碱解氮含量显著降低。滇黄精是多年生宿根草本植物,以地下根状茎为主要收获对象。在本研究中,自2018年起开始用滇黄精进行间作试验,其生长阶段对速效钾、有效磷、碱解氮等养分有不同的需求,并且会将大量养分向根茎积累[20],导致土壤养分含量下降。这一现象与严毅等[10]的研究结果相吻合,他们发现在油橄榄林下种植百合后,土壤中营养成分含量均明显降低,表明百合生长对有机质、氮、磷、钾等养分有不同程度的吸收需求。因此,在间作滇黄精期间,应注意其与油橄榄之间的养分竞争,注意加强施肥管理。FZ间作模式显著增加了土壤的全钾、有效磷含量,但有机碳、碱解氮、速效钾含量在间作后显著降低。紫花苜蓿草为深根性多年生牧草,有研究结果表明豆科作物具有较强的质子释放能力,其根系对土壤中难溶性磷的活化和其他作物吸收磷有促进作用[21-22],进而提高了作物的磷含量。大量生产实践结果表明,紫花苜蓿草的根瘤有固氮的作用[23-24],可在一定程度上增加土壤中氮含量。本研究中可能由于紫花苜蓿草间作年限较长,其根系会分泌一些不易降解的酚酸类物质,这些物质累积在土壤中影响其生长[25],从而影响养分吸收与转化,同时间作后紫花苜蓿草与油橄榄树存在养分争夺的现象,导致土壤中氮含量显著降低。F3间作模式下土壤全钾含量显著增加,除碱解氮外其他养分含量相较于CK均有所增加,其原因是多种牧草根系在地下的分布存在生态位差异[26],可充分协调水、养、气的协同[27],这与王雪等[28]、姜黎等[29]、张永亮等[30]的研究结果相似,同时混种间作作物占据了更多生态位,增加了群落的稳定性,其凋落物累积量也会增加,从而回归土壤的养分增多[31]。
3.2间作模式对土壤微生物多样性和群落组成的影响
间作对土壤微生物多样性的改变通常与土壤环境、作物种类、间作年限等多重因素相关。OTU分析与α多样性分析结果显示,间作后细菌和真菌多样性指数无显著差异,但F3间作模式在一定程度上增加了土壤中细菌、真菌的OTUs数量,这也反映了多草种间作有利于提高土壤微生物多样性,3种间作处理下细菌群落物种组成与CK相似,均以放线菌门、变形菌门、酸杆菌门为优势细菌菌门,这些菌门在油橄榄土壤中的优势地位已有报道[32],但间作后各菌群的相对丰度有一定变化,由于不同植物的根系分泌物和凋落物理化性质不同,其分解时释放的养分具有差异,进而导致各处理中优势菌门的相对丰度存在差异[33-34]。子囊菌门、担子菌门、被孢霉门为真菌优势菌门,这与许彦明等[35]的研究结果一致。子囊菌门作为真菌界种类最多的门,在土壤有机质的降解中具有重要作用[36]。真菌门水平上,3种间作处理相较于CK均增加了子囊菌门的相对丰度,这可能与间作作物的分泌物、凋落物为其提供营养与能量有关。被孢霉门具有溶磷性[37],被孢霉门为FZ间作模式下土壤的优势菌门,这与FZ间作模式显著增加了土壤中有效磷含量的分析结果一致。在真菌属水平上,各模式间优势属组成及相对丰度的差异均较大,间作均增加了其他功能真菌群落的相对丰度。其中间作滇黄精(FD)和紫花苜蓿草(FZ)后镰刀菌属的相对丰度明显增加,镰刀菌属为常见的根腐病病原菌,并在多种植物中被检出[38-39],因此间作滇黄精与紫花苜蓿草后可能出现病害侵扰。相较于CK,F3模式下土壤中古根菌属、青霉属、小囊菌属等有益微生物的相对丰度明显增加,这种间作模式有利于提高土壤有益功能真菌的丰富度,多草种间作通常对土壤温度、养分等土壤环境因子有明显影响,从而对微生物群落种类和结构产生直接或间接影响[40],导致群落组成结构产生差异。
3.3土壤微生物群落组成与土壤环境因子的关系
土壤微生物对环境极为敏感,土壤类型和理化性质的改变会引起土壤微生物群落组成发生变化,主要菌群的丰度和结构也不同[41]。土壤理化性质对土壤微生物群落的影响较为复杂,目前尚无完全一致的结论。颜彩缤等[42]通过对3种菠萝蜜间作模式进行研究,发现放线菌门与土壤全钾、全氮、全磷含量均呈现极显著或显著负相关;何升然等[43]通过对紫花苜蓿与甜高粱间作进行研究,发现酸杆菌门和疣微菌门与土壤pH值呈现极显著正相关,与土壤碱解氮含量呈现极显著负相关。李敏等[44]发现氮和磷含量能够显著影响根际土壤真菌群落,氮元素的转化和循环与土壤真菌有着密切的关系,磷元素在真菌的繁殖和生长中也发挥了重要的作用。相关性分析结果表明,土壤有机碳、全氮、全磷、全钾、速效钾、碱解氮、有效磷的含量与部分细菌门的相对丰度有极显著相关性,土壤有机碳、全氮、速效钾、碱解氮的含量与部分真菌门的相对丰度有极显著相关性,表明养分元素对这些微生物的生长和代谢具有促进或抑制作用,间作通过改善土壤理化性质为土壤部分菌群提供了适宜的营养环境,进而使得土壤微生物数量、活性及群落组成发生变化[45-46],李承臻等[47]的研究结果表明,在荔枝园间作柱花草可以改变土壤微生物群落结构,微生物群落相对丰度的变化与间作柱花草后氮、磷、钾等土壤养分含量的变化有关,这与本研究结果一致。同时部分养分含量与某些细菌、真菌存在抑制效应,土壤中某些养分过量可能抑制这些微生物的生长或导致其处于竞争劣势[48]。
油橄榄×滇黄精(FD)、油橄榄×紫花苜蓿草(FZ)、油橄榄×紫花苜蓿草×黑麦草×金鸡菊(F3)3种间作模式实施5a后,土壤化学性质均发生了改变。FD模式和FZ模式间作后土壤中部分养分含量显著降低,间作作物与油橄榄存在养分争夺现象,F3模式提高油橄榄林地土壤养分含量的效果最佳。3种间作模式对土壤微生物多样性影响较小,但可诱导土壤细菌、真菌群落结构发生变化,以真菌群落结构的变化最为明显。3种间作模式均明显增加了部分土壤真菌有益微生物的相对丰度,但FD和FZ模式增加了常见部分土壤致病菌属的相对丰度,F3间作模式对维持和促进土壤生态系统良性发展具有积极影响。土壤细菌与真菌群落结构变化受土壤化学性质驱动,土壤养分含量对部分细菌和真菌群落组成和丰度有重要影响。综上,油橄榄×紫花苜蓿草×黑麦草×金鸡菊(F3)多草种间作模式优于油橄榄×滇黄精(FD)和油橄榄×紫花苜蓿草(FZ)间作模式。本研究中仅测定分析了间作模式下油橄榄林地表层土壤养分含量指标和土壤微生物多样性及群落结构变化的差异,未涉及不同深度土壤的养分含量和微生物群落变化,后续将进一步研究间作对油橄榄林地不同土层微生物群落和养分变化的影响,同时间作作物对油橄榄树养分吸收及果实产量和品质的影响程度也有待进一步探究。
