贵州豹皮樟叶表型与解剖结构特性及其与环境因子关系
2024-12-27廖梦芸袁刚毅谢娜高超赵彤郭其强


关键词:豹皮樟叶片;表观参数;解剖结构参数;环境因子
叶片是植物进行能量转化和生物量积累的重要器官,其表型特性和功能受生境因子影响显著[1]。叶片表型与解剖结构作为叶片重要功能性状,不但能感受生境因子变化并做出响应,而且能长期积累该响应后表现出特有的生存能力和适应策略[2-3]。因此,探究植物叶片表型特性及其与环境因子的关系不仅能明确其适生特性,而且对于特有物种资源保护及定向种质筛选具有重要意义。
植物叶片表观特性及解剖结构因生境地环境因子不同而表现出特定变异规律,即叶表型变异。例如,干旱条件下,植物叶面积趋于变小,栅栏组织加厚而海绵组织变薄;当降水量增加时,植物比叶面积则趋于增加[4]。高温环境下叶片细胞质壁分离不明显,叶肉细胞排列紧密;而低温下叶片厚度、输导组织和维管束厚度增加,使得叶片厚度及干物质量均较高[5]。此外,环境中降水量及其季节分布格局的差异也会导致植物叶片功能性状的改变[6-7]。上述现象是植物对环境因子变化做出的有利响应,并使该物种得以长期生存[8]。因此,阐释种群间和种群内植物叶片变异特性对于明确物种在异质生境下的生存策略及其可塑性具有重要价值。
豹皮樟Listacoreanavar.sinensis是樟科Lauraceae木姜子属Litsea常绿小乔木,天然分布区主要位于我国西南部。叶片可制茶,饮用历史悠久[9];其叶片富含黄酮、蛋白质、氨基酸等次生代谢物,有解毒、消肿、抗炎及抗氧化特性,极具应用价值[10-11]。贵州省是豹皮樟资源主要分布区之一,目前因过度采摘,种群生长势差,无法结实及幼苗更新,处于濒危状态[12],亟需加强保护。以往学者们对豹皮樟的研究主要集中在资源分布及利用价值等方面[9-13],而对豹皮樟叶器官生物学特性研究较少。因此,有关豹皮樟在异质生境的生存偏好及表型特性仍不清楚,导致无法确定有针对性的保护措施。本研究拟采集贵州省不同分布区豹皮樟成年植株叶片,通过石蜡切片观察和相关分析探讨不同豹皮樟种群叶片表型特性及其与环境因子的关系,为进一步明确该物种生存特性及实施有效的保护与管理提供科学依据。
1材料与方法
1.1研究区概况
研究区位于贵州省5个县域,地形起伏大且地貌类型多样,海拔高度为650~870m。植被以马尾松林Pinusmassoniana、常绿阔叶林和竹林等类型为主。该区域属亚热带湿润季风气候,雨热月份相同;年均气温约15.0℃,平均气温1月最低,约1.0℃,而7月最高,约28.0℃;年均降水量1255mm,无霜期约为300d;土壤以山地黄壤为主。
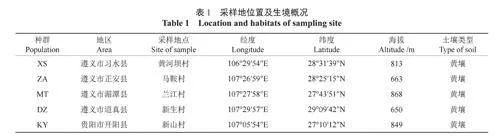
根据前期踏查贵州省豹皮樟资源分布情况,本研究选择5个豹皮樟分布较为集中的县进行采样,分别为遵义市的习水县、正安县、湄潭县、道真县和贵阳市的开阳县。在这5个县采集的种群分别命名为习水种群(简称:XS)、正安种群(简称:ZA)、湄潭种群(简称:MT)、道真种群(简称:DZ)和开阳种群(简称:KY)。材料采集地位置及生境概况见表1。
1.2样品采集
在每个种群选取生长良好的成年个体20株,在每个样株上按树冠的东、南、西、北向采集完整、生长正常的当年生叶40片,均分为2份。1份放入保鲜袋带回实验室用于表型测定;1份立刻放入FAA固定液中保存,带回实验室制作切片。
1.3试验方法
叶片表观参数测定:使用叶面积仪(AM350)测定叶长、叶宽及叶面积;使用精度为0.01mm的游标卡尺测定叶柄长;使用SPAD502叶绿素仪测定叶绿素值;用精度为0.01g电子天平测定单叶鲜质量,后将其于80℃烘箱烘干至恒定质量,测量其干质量[14]。同步计算叶形指数、叶干物质量及比叶面积等参数。
叶片解剖结构参数观测与分析:剪取叶片中部横切部分,经酒精脱水,用二甲苯透明处理后石蜡包埋、切片,染色和脱水处理后用树胶封片,放于光学显微镜(LeicaDM2500)下拍照保存。利用ImageJ软件观察和测量叶片上角质层厚度、上表皮厚度、栅栏组织厚度、海绵组织厚度、下表皮厚度、下角质层厚度和主脉厚度[14],并计算栅海比(栅栏组织厚度/海绵组织厚度)。
1.4环境因子数据获取
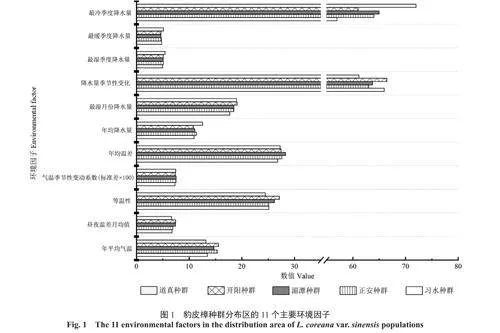
气候作为重要环境因子,数据来源于WorldClimV2.0(http://www.worldclim.org)。根据采样点地理信息,利用R语言(4.3)Raster软件包对植物采样点对应环境因子变量进行提取,共获取19个环境因子变量(1970—2000年),去除各强相关性因子(r2≥0.9)后,最终确定11个相对独立的变量因子[14],分别为年平均气温,昼夜温差月均值,等温性,气温季节性变动系数,年均温差,年均降水量,最湿月份降水量,降水量季节性,最湿季度降水量,最暖季度降水量,最冷季度降水量(图1)。
1.5数据处理与分析
采用SPSS22.0软件进行数据统计,计算各参数变异系数(CV),使用嵌套式方差分析计算叶表型分化系数(Vst)[14]。计算公式为:
使用Origin2024b进行RDA计算并绘图,筛选影响叶表型和解剖结构参数的主要环境因子。
2结果与分析
2.1叶片表观特性参数及变异性分析
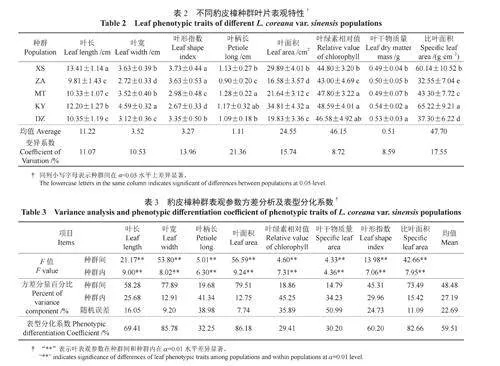
统计豹皮樟不同种群叶片表观特性参数(表2),发现各种群间叶片表观特性参数均存在极显著的差异性。如,XS种群和KY种群叶片宽大,而XS种群叶呈狭长形且干物质量最低,而KY种群叶片较宽阔;ZA种群叶片较小,且比叶面积与叶绿素相对值均较低;MT及DZ种群叶片表观参数居于中等,但DZ种群较MT种群叶片更狭长。豹皮樟叶片表型参数的变异系数介于8.59%~17.55%,说明豹皮樟种群具有一定的变异和表型多样性。
由表3可知,豹皮樟种群叶片表观参数在种群间和种群内均存在极显著差异。种群间和种群内的方差变异分量分别占总变异量的48.48%和27.19%;表型分化系数的变化区间为29.41%~86.18%,其中叶面积(86.18%)、叶宽(85.78%)和比叶面积(82.66%)分化系数较大,而叶绿素相对值(29.41%)、叶干物质量(30.20%)和叶柄长(32.35%)分化系数较小。豹皮樟叶片表观性状在种群间表型分化系数为59.51%,说明豹皮樟叶片表观性状的变异约60%来自种群间。由此可见,豹皮樟种群间的变异是造成该物种叶片表观参数变异的主要因素。
2.2叶片解剖结构特性及其变异性分析
豹皮樟叶片为异面叶(图2),其解剖结构由角质层、表皮、栅栏组织和海绵组织组成,叶脉结构完整;栅栏组织细胞排列紧密,而海绵组织细胞稀疏且厚度较大[15]。
由表4可知,不同种群叶片解剖结构参数均值中,KY种群叶片解剖结构较发达,除下表皮细胞厚度值外,其余各参数均最大;而XS种群下角质层厚度和栅海比,ZA种群上角质层厚度、栅栏组织厚度和主脉厚度,MT种群海绵组织厚度和DZ种群的上表皮细胞厚度、下表皮细胞厚度值均最小。豹皮樟各解剖结构参数变异系数为6.70%~16.81%,其中下角质层厚度变异系数最大,主脉厚度变异系数最小。
由表5可知,在种群间和种群内8个解剖结构参数均差异极显著(P<0.01);各参数在种群间方差分量占总变异的57.92%,种群内占比为23.83%;表型分化系数变化幅度为49.28%~89.60%,其中下表皮厚度(89.60%)和上角质层厚度(88.99%)的表型分化系数较大,而下角质层厚度(49.28%)、主脉厚度(58.60%)和栅海比(58.31%)的表型分化系数较小。各参数种群间平均表型分化系数为69.55%,说明豹皮樟叶片解剖结构参数的变异约70%来自种群间。该结果表明叶片解剖结构参数变异主要来源于种群间。
2.3叶片表型和解剖结构与环境因子关系分析
豹皮樟叶表型和解剖结构与环境因子间存在显著相关性。由图3(a)可知,第一主成分和第二主成分两个主成分能解释环境因子对豹皮樟叶表型参数影响力的82.70%,其中叶表型参数中叶长、比叶面积、叶面积和叶宽与降水量季节性变化、年均降水量、等温性等环境因子间存在显著正相关关系,而与气温季节性变动系数和最冷季度降水量等环境因子间存在显著负相关关系。由图3(b)可知,第一主成分和第二主成分两个主成分能解释环境因子对豹皮樟叶片解剖结构参数影响力的81.56%,其中下表皮厚度、上表皮厚度、下角质层厚度、栅海比等解剖结构参数与降水量季节性变化存在显著正相关关系,而与气温季节性变动系数、年均温差间呈显著负相关关系。
3讨论
3.1叶片表观参数变异性度及成因分析
植物天然种群表观参数变异性分析不仅反映了其变异度和变异来源,而且可作为资源鉴定、保存、筛选与利用的基本依据[16-17]。豹皮樟叶片表观参数在种群间和种群内均为极显著差异,说明豹皮樟在种群内和种群间均有高度特异性。该特性与火棘Pyracanthafortuneana[18]变异特性类似。从分化程度看,豹皮樟平均分化系数为62.36%,高于格木Eredphleumfordii(21.86%)[19],但低于云南黄连Coptisteeta(74.41%)[20]和夏蜡梅Sinocalycanthuschinensis(89.30%)[21]。综上来看,贵州豹皮樟种群叶片表观参数总体处于中等分化水平,也具有明显的环境选择性。
植物叶片表观参数是在长期进化和生境适应中逐渐积累的结果。叶面积和长宽比会影响植物对光合有效辐射的接受能力以应对温度变化[22];比叶面积代表植物从外界获取碳的能力[23];叶柄长度能够让植物改变叶片伸展度以调节受光量[22],从而控制光合效率;叶绿素含量决定植物光能转换的能力,是叶片光能利用效率的最直接因素;而叶干物质量代表植物利用资源的能力及对不利生境的适应性[24]。本研究发现,豹皮樟叶片叶长、叶宽、叶形指数、叶面积和比叶面积为高度分化(Vst>60%),仅有叶柄长、叶绿素相对值和叶干物质量为低度分化(Vst<40%)。这说明该物种叶片具有显著的表型可塑性,该特性使得物种生存偏好特定的生境。已有研究发现,豹皮樟主要喜荫蔽生境[9,12],造成叶柄长和叶绿素低分化度低。然而,植物偏好特定生境对于该物种分布区扩张和应对分布区生境改变是极其不利的。实际踏查中发现,野生豹皮樟幼苗一旦被村民发现,就会被移栽至自家房前屋后,这极易造成幼苗死亡,这也是造成豹皮樟野生资源濒危的重要因素之一。
豹皮樟叶片表观参数变异首先源自种群间,其次为种群内,这与云南松P.yunnanensis表观多样性变异特性一致[25]。一方面说明豹皮樟对环境压力表现出一定的适应性,即种群间表型分化水平与种群对环境的适应能力成正比;另一方面与物种繁殖特性有关[14]。分析原因如下:研究区属于高原山地,多变的气候和复杂的生境条件使得豹皮樟形成了特定的适应性并得以长期维持。同时,由于生境区破碎化程度高,使得各种群间基因交流困难,容易造成地理和生殖上的隔离[26-27],由此导致种群间表观变异较高。
3.2叶片解剖结构参数变异性来源分析
植物叶片解剖结构参数因生境不同而导致显著差异分化[28]。豹皮樟叶片解剖结构参数在种群间和种群内均呈极显著差异,其种群间的方差分量为57.92%。这不仅表明其变异主要来源自种群间,而且进一步说明了豹皮樟叶片对环境因子变化较敏感。在不同的生境条件下形成了特有的适应类型,并最终表现为解剖结构参数的差异性。因此,研究豹皮樟叶片解剖结构特征也可为野生资源收集、良种选育等提供重要参考[29]。
表型变异系数不但反映植物叶片表型性状的变异程度,而且反映资源表型的丰富度[30]。本研究中豹皮樟叶解剖结构参数平均变异幅度为6.70%~16.81%,与板栗(9.81%~15.60%)和闽楠(3.73%~32.28%)的变异系数值差异不大[31]。这说明豹皮樟叶片解剖结构各参数总变异度较低。学者们研究发现,变异系数值也是植物表型结构的生态适应参数;若变异系数小于50%,则认为其表现结构相对稳定,相反则为不稳定[32]。综上可知,豹皮樟叶片解剖结构参数虽有一定变异性和丰富度,但总体仍为稳定型。该特性对于维持物种自身生理稳定是有利的,这也是植物适应复杂生境的重要生存策略之一。
3.3叶片表型参数与环境因子的关系分析
叶片表观参数是基因和环境互作的结果[33],物种在不同环境中因适应机制和环境敏感性的差异形成了特定的地理变异规律[27-28]。本研究发现,豹皮樟叶片表型参数中叶长、叶宽、叶面积和比叶面积与环境因子中的等温性、年均降水量和降水量季节性变化呈显著正相关,而与气温季节性变动系数和最冷季度降水量间存在显著负相关。这说明年均气温波动小,气候温暖、雨水充沛的气候条件更利于豹皮樟叶片发育,并形成宽大的叶片,而明显的季节性温度变化和冷季降雨对叶片发育是不利的。曾维军等[34]研究发现,豹皮樟碳光合作用与环境湿度和温度关系密切,而干旱条件下其净光合速率会显著降低。这也说明适宜的水分和温度条件有助于豹皮樟叶片维持较高光合效率,积累大量的营养物质来促进叶原基细胞的分裂和数量增加,使得叶边缘和顶端快速生长,最终形成宽阔的叶片[35]。因此,本研究中XS和KY种群发达的叶表型参数与其生境区气候条件密切相关。由此可见,生境区温暖、湿润的气候条件更有利于豹皮樟形成宽大的叶片。
植物叶片微观结构与环境因子关系密切,后者通过影响叶片微观结构导致叶片厚度分异[36]。本研究中,豹皮樟下表皮厚度、上表皮厚度、下角质层厚度、栅海比等解剖结构参数与降水量季节性变化存在显著正相关,而与气温季节性变动系数、年均温差呈显著负相关。这说明季节性降水充足有利于豹皮樟叶表皮细胞发育,同时形成发达的栅栏组织,导致叶片厚度增加;而气温大幅度波动,如气温的季节性变化和年均温度较大,则会阻碍叶肉组织发育。在本研究中,豹皮樟生境地均为亚热带气候区,降雨多集中在4—6月份,此时正值植物叶片由幼嫩转向成熟的过渡期,充足的水分和相对较高的气温极有利于植物细胞增殖,从而形成厚实的叶片。此外,由于栅栏组织含有丰富的叶绿体,能促进植物高效吸收和转化光能,完成能量转化和养分积累[33],这对于植物更好利用光能和适应荫生环境都具有积极意义。
4结论
豹皮樟习水和开阳种群叶片较宽大;叶片表观参数变异度为种群间高于种群内;叶片解剖结构为异面叶,其中开阳种群解剖结构较发达。环境因子中,夏季降水和一定的温度条件是影响豹皮樟叶片表型和解剖结构参数的主要环境因子。开展资源保护时,建议首选开阳种源;人工栽培时应选择降水充沛且气温较高的区域,以利于豹皮樟叶片发育和植株生长。本研究阐明了贵州豹皮樟种群叶片表型基本特性,相关结论可为有效保护豹皮樟资源及高效种植提供一定科学依据。但是,由于研究区环境因子多样性和气候因素复杂性,有关豹皮樟生理特性研究仍需进一步深入和完善。
