氮添加对水杉防护林生长的影响
2024-11-03姚洪锡冯坤乔童冉原文文吴统贵朱念福





摘要:[目的]在我国大气氮沉降量逐年增加背景下,厘清外源氮素输入对水杉防护林生长的影响,有助于为防护林健康生长开展养分管理提供科学指导。[方法]以江苏省东台市泥质海岸12年生水杉(Metasequoiaglyptostroboides)防护林为研究对象,通过设置4个水平的连续8a氮添加处理,即对照(CK,0 kg·hm-2·a-1)、低氮(LN,56 kg·hm-2·a-1)、中氮(MN,168 kg·hm-2·a-1)和高氮(HN,280 kg.hM·-2·a-1),研究氮添加对水杉及其大小树(根据初始平均胸径划分)胸径年生长量的影响。[结果]氮添加水平和年份及其交互作用显著影响水杉胸径年生长量。氮添加对水杉胸径年生长量呈非线性影响,中氮添加的促进作用强于低氮和高氮。年际间水杉胸径生长量的变化趋势一致:初期氮添加促进作用逐渐增强(1-3年氮添加),然后促进逐渐减弱(4-6年氮添加),继而转变为抑制作用(7-8年氮添加)。在同水平氮添加处理下,径级和年份显著影响水杉生长,但无交互作用。对于小树,氮添加水平和年份及其交互作用显著影响胸径生长。对于大树,仅氮添加年份显著影响胸径生长。水杉小树的胸径年生长量低于大树,而生长量响应比则高于大树。[结论]同水平氮添加处理下,水杉大树胸径生长量高于小树,但氮添加对小树胸径生长的促进作用强于大树,表明小树生长更易受外源氮输入的调控。同时,随着年份增加,氮添加对水杉胸径生长表现出先促进后抑制,表明长期氮沉降将不利于泥质海岸水杉防护林生长。
关键词:氮添加;水杉防护林;胸径年生长量;小树;大树
中图分类号:S750 文献标识码:A 文章编号:1001-1498(2024)05-0186-09
我国目前已经成为世界上活性氮(N)排放量最多的国家,其中东部沿海地区是最严重的区域之一,大于30 kg·hm-1·a-1,对该区域森林生态系统产生了深远影响。树木作为森林生态系统的骨架,是森林生态系统物质循环、能量传递的重要载体,其生长状况在氮沉降背景下的变化,受到广泛关注。
已有研究表明,氮沉降对树木生长的影响存在3种结论:促进作用、抑制作用、无显著影响。日渐丰富和完善的长期N添加和联网试验,以及文献综述和荟萃分析(Meta-analysis)研究,发现气候、林龄、植物功能性状、径级等因素参与调控外源性氮输入对树木生长的影响。
目前氮添加对树木生长的影响研究中,大部分以林分尺度的整体生长量(生长率)或生物量为对象,而忽视了树木大小的调控作用。有研究表明在亚热带和温带的一些区域,氮添加后大树生长要快于小树,但亦有研究表明小树生长较快,可能与养分限制元素、树种和功能性状等因素有关。在同一林分中,小树对氮添加的响应具有不确定性,既可能是促进作用,也可能是抑制作用,这种矛盾的结论可能与小树和大树之间的资源竞争类型和强度有关。氮添加对树木生长的影响在持续时间上也存在差异,已有研究表明短期氮添加将显著促进树木生长,随着施氮年限的延续,促进作用逐渐减弱,甚至转变为无明显作用或抑制作用。
水杉(Metasequoia glyptostroboides Hu&W.C.Cheng)适应能力强,耐盐耐寒,喜湿润,生长快速,抗风能力强,是我国东部泥质海岸防护林第一大造林树种,造林面积600多万亩,占该区域基干林带总面积的50%以上:201,具有良好的防护效益。因此研究氮沉降对水杉防护林生长的影响将有助于为防护林质量提升开展养分管理提供科学指导,具有重要的实践价值。本研究以江苏省东台市泥质海岸水杉防护林为研究对象,通过4个水平的连续8a氮添加试验,以期完成以下研究目标:1)不同水平氮添加和年份对水杉生长有何影响?2)不同大小的水杉生长对氮添加的响应有何差异?
1 材料与方法
1.1 样地概况
试验区域位于江苏省东台市黄海海滨国家森林公园(120°07'~120°53'E,32°33'~32°57'N),是华东地区规模最大的人造生态林园,距离海岸带直线距离约14公里,地貌属苏北海洋堆积平原,气候属于亚热带和暖温带的过渡区域,热量充裕,雨量充沛。年均气温15℃,年均降雨量1 061 mm,年均相对湿度88.3%,年均无霜期22d,年均日照时长2 130 h。水杉为该区域防护林主要树种之一。
1.2 试验设计
2014年,在试验区域选取3个立地条件和营建方式(林分密度等)基本一致的12年生水杉防护林林分,在每个林分中选定一块300 m × 100 m的区域作为样地。在每个样地中建立4个30mx20 m的样方,样方间隔大于10 m,设置4个氮添加处理,每个处理3个重复,3个林分共计12个试验小区。参考该区域同期大气氮沉降量,以及同类研究通用的施氮量成倍增加原则,设置4个水平氮添加处理:对照(CK,0 kg·hm-2·a-1)、低氮(LN,56 kg·hm-2·a-1)、中氮(MN,168 kg·hm-2·a-1)、高氮(HN,280 kg·hm-2·a-1)。
在2015年,使用尿素(CO(NH2)2)分两次在水杉样方内均匀撒施,第一次在3月底至4月初,按总量的60%进行添加;第二次在6月中旬,按总量的40%进行添加,此后每年均采用该方式进行施肥。
1.3 树木测定
2015年施肥之前,对样方内的水杉进行每木检尺,记录胸径、树高、冠幅和枝下高等信息,并进行编号,之后每年在同一时间段进行树木测定。其中2020年进行了施肥,但未进行树木测定,因此2020年对应的胸径年生长量为2019年到2021年水杉胸径生长量的一半。当试验期间有树木死亡、折断、倒伏等,记录其编号,但不进行处理,样方基本信息如表1所示。
1.4 数据分析
根据2015年各样方胸径平均值划分大小树,胸径大于平均值为大树,小于平均值为小树。采用胸径年生长量表示水杉生长对N添加的响应,胸径年生长量公式为:
△DBH=DBHt2-DBHt1
式中,△DBH为胸径生长量,DBHt2为本年度的胸径值,DBHt1为上一年度的胸径值。
利用自然对数转换的胸径年生长量响应比(InRR)评估水杉生长对不同水平氮添加的响应变化趋势,响应比公式为:
In RR=ln(Xt/Xc)
式中,Xt为氮添加处理(LN、MN或HN)的胸径年生长量,Xc为同期对照处理的胸径年生长量。
利用双因素方差分析(Two-way ANOVA),计算水杉胸径年生长量在不同氮添加水平和不同年份之间的差异。以上统计显著性水平设置为P=0.05,数据分析和绘图均采用R软件(version4.3.1)。
2 结果与分析
2.1 水杉胸径年生长量对氮添加的响应
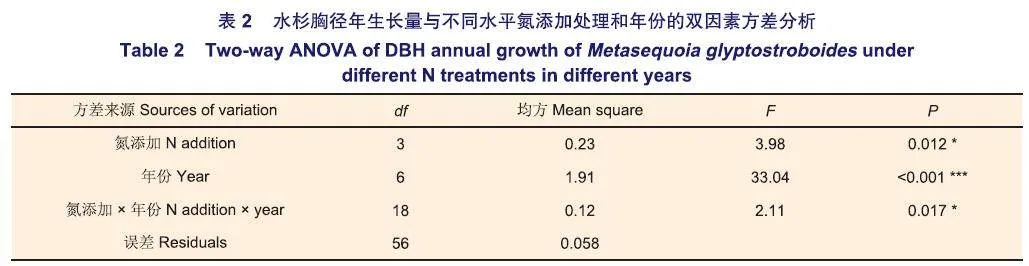
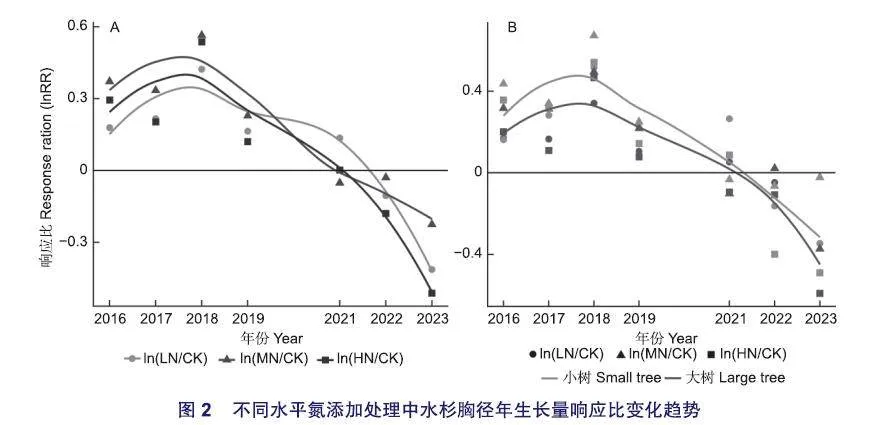
水杉胸径年生长量对不同水平氮添加和年份及其交互作用的响应存在显著差异(表2,图1)。不同水平氮添加明显改变了水杉胸径生长过程,MN处理对水杉胸径年生长量的促进作用强于LN和HN(图1,图2A)。从2016年到2023年,各水平氮添加下胸径年生长量均呈“减少一增加一减少”变化趋势(图1),CK处理中最大年生长量出现在2022年,LN、MN和HN处理中最大年生长量出现在2016年。
从2016到2018年(1-3年氮添加),LN、MN和HN处理均增加了水杉胸径年生长量(图1),且促进作用逐渐增强(图2A);其中LN处理均无统计学差异,HN处理在2016年和2018年时促进作用显著,而MN处理则在2016年和2018年均显著促进水杉生长(图1)。从2019年到2021年(4-6年氮添加),LN、MN和HN处理对水杉胸径年生长量的促进作用逐渐减弱(图2A),各水平氮添加均无统计学差异。从2022年到2023年(7-8年氮添加),LN、MN和HN处理减少了水杉胸径年生长量,变成抑制作用(图1,图2A);其中LN和MN处理无统计学差异,HN处理在2023年抑制作用显著。
在CK处理中,2018年胸径年生长量最小,显著低于其它年份;且2021年胸径年生长量显著低于2016年、2022年和2023年。在LN处理中,2023年胸径年生长量最小,显著低于2016年。在MN处理中,2021年胸径年生长量最小,显著低于2016年、2017年和2022年;且2018年、2019年、2022年和2023年的胸径年生长量显著低于2016年。在HN处理中,2023年胸径年生长量最小,显著低于2016年和2017年;且2018年、2019年、2021年和2022年的胸径年生长量显著低于2016年。
2.2 不同大小水杉胸径年生长量对氮添加的响应
同一水平氮添加中,径级和年份对胸径年生长量有显著影响,但交互作用不显著(表3)。各水平氮添加中,小树和大树胸径年生长量均呈“减少一增加一减少”变化趋势(图3)。在CK处理中,2016年、2017年和2023年小树胸径年生长量显著低于大树,其中2018年小树和大树生长量均最小。在LN处理中,仅2016年小树生长量显著低于大树,且2023年小树和大树生长量均最小。在MN处理中,各年份小树生长量与大树无统计学差异,其中2021年小树和大树生长量均最小。在HN处理中,仅在2022年的小树生长量显著低于大树,其中2023年小树和大树生长量均最小。
对于小树,氮添加水平和年份对胸径年生长量具有显著影响,且交互作用显著(表4)。从2016年到2019年(1-4年氮添加),MN处理显著促进水杉胸径生长,从2016年到2017年(1-2年氮添加),HN处理显著促进水杉胸径生长;2023年(8a氮添加)HN处理显著抑制了水杉胸径生长。
对于大树,年份对胸径生长量具有显著影响,而氮添加水平及其交互作用影响不显著(表4)。2016年(1a氮添加),MN处理显著促进水杉胸径生长;2023年(8a氮添加),MN处理抑制了水杉胸径生长,但无统计学差异。2023年(8a氮添加),HN处理显著抑制了水杉胸径生长。同时,连续8a LN处理对小树和大树胸径年生长量差异均无显著影响。
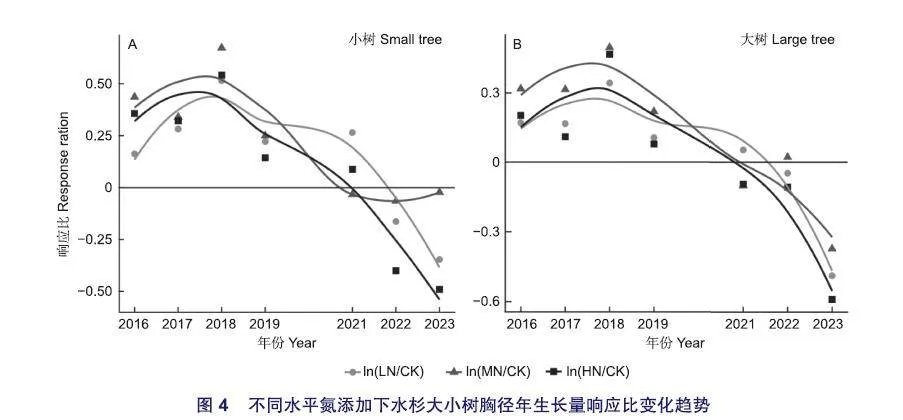
同一水平氮添加处理中,小树胸径年生长量响应比均高于大树(图2B,图4),但变化趋势基本一致,即随氮添加年份增加呈先增加后减少变化。从2016年到2018年响应比逐渐增加,在2018年达到峰值,然后从2019年到2021年响应比逐渐减小(大于0),从2022年到2023年响应比继续减小(小于0)。
3 讨论
3.1 水杉胸径生长对氮添加的响应及年际间变化
传统研究观点认为,在北半球纬度梯度上热带森林生长主要受磷限制,北方森林主要受氮限制,过渡区域从磷限制逐渐转变为氮限制。本研究结果表明,短期氮添加(1-6年氮添加)增加了水杉的胸径年生长量(图1),说明氮素是该区域限制水杉生长的一个重要因素,与传统森林生态系统元素限制理论预测结果一致。
多数研究表明树木生长与氮添加水平呈非线性关系,表现为中氮促进作用强于低氮和高氮,本研究结果同样支持该结论(图1,图2A),原因可能是低氮添加不能满足养分需求,而高氮添加则暂时可能超过养分需求,导致化学计量比失衡抑制生长。低水平氮添加在短期内(1-3年氮添加)对水杉胸径年生长量无显著影响,而中氮和高氮则显著增加了生长量,与刘修元等对落叶松[Larix gmelinii (Rupr.) Kuzen.]原始林连续3a氮添加促进胸径生长的研究结论一致,原因可能是低水平氮添加对树木的影响受到林下植被的抑制,或被树木优先用于改变功能性状,如增加叶片氮含量。随着氮添加年份增加(4-6年氮添加),各水平氮添加处理对胸径年生长量无显著影响,这与Kjonaas等在瑞典针叶林开展持续5a氮添加对树木生长速率无显著影响结论一致,原因可能是前期持续氮添加导致生态系统接近氮饱和,以及较高的环境氮沉降速率能够满足树木生长需求,从而对额外氮添加无明显响应。若继续添加氮可能不利于树木生长,而本研究中随着各水平氮添加处理继续进行(7-8年氮添加),水杉胸径年生长量小于对照处理证明了这一点,可能原因是持续外源氮输入超过了植物对氮的同化作用,打破了植物体内化学计量耦合关系的平衡,从而表现为抑制作用。
从2016年至2023年(1-8年氮添加),水杉胸径年生长量表现出“减少-增加-减少”的复杂变化趋势,可能与年际间气候条件(温度、降水等)剧烈变化有关,对树木生长产生复杂的交互影响;或者是长期氮添加造成前期氮积累,从而对后期树木生长产生影响,表现出氮滞留效应;亦或是随着林龄增加,树木生长所需养分随其发育阶段变化,从而对氮添加的响应发生了变化。
3.2 不同大小水杉胸径生长对氮添加的响应
已有研究发现氮添加对树木生长的影响因树木径级的不同而存在差异。本研究结果表明在相同水平氮添加处理下,水杉小树胸径年生长量低于大树(图3),与同类研究结论一致,即氮添加对树木生长的影响与树木的初始大小相关,原因可能是大树对养分的获取能力强于小树。
水杉小树和大树胸径年生长量响应比随氮添加年份增加的变化趋势为促进作用先增强,然后逐渐减弱,最后变为抑制作用,可能与植物对土壤氮的周期性响应有关,或持续氮添加可能影响微生物、土壤和叶片化学计量平衡,从而改变了养分限制状况。同水平氮添加处理下,水杉小树胸径年生长量响应比高于大树(图2B),可能是由于小树吸收养分既要用于促进植物生长,又要用于增加叶片养分含量以提高光合效率,因此更易受养分限制,说明水杉小树胸径生长更容易受到外源氮输入的影响。
4 结论
以江苏省东台市泥质海岸12年生水杉防护林为研究对象,通过4个水平的连续8a氮添加试验,研究发现:
(1)氮添加对水杉胸径年生长量的影响呈非线性变化,中氮添加促进作用强于低氮和高氮。随着氮添加年份增加,较对照处理,各水平氮添加处理影响趋势一致:促进作用逐渐增强(1-3年氮添加),然后促进作用逐渐减弱(4-6年氮添加),最后变为抑制作用(7-8年氮添加),表明长期氮沉降将不利于水杉防护林生长。
(2)同水平氮添加处理下,水杉小树胸径年生长量小于大树,但胸径年生长量响应比高于大树,表明外源氮输入对小树生长的促进作用强于大树,小树生长更易受外源氮输入的调控。
(责任编辑:崔贝)
基金项目:浙江省科技计划项目(2022C02053);国家自然科学基金项目(33201632,31770756)
