鱲属鱼类研究进展
2024-10-27席红波宋雪琳

摘要:鱲属目前仅有4个已定名的有效种,即宽鳍鱲、棘颊鱲、中华鱲和苕溪鱲。本文查阅了国内外相关文献,对鱲属鱼类的生态学(生活习性、食性、年龄组成与生长及繁殖)、遗传多样性、系统发育及资源现状等方面的研究进展予以综述,并就资源保护与研究提出建议。
关键词:鱲属;分类;研究进展;系统发育
中图分类号:S917.4 文献标识码:A
基金项目:2021年江西省教育厅科学技术研究课题“台湾白甲鱼物种复合体的分类界定、物种隐存多样性与分子系统地理结构分析”(GJJ211133)
作者简介:席红波(2002—),男,本科,研究方向:生物科学。E-mail:3114962697@qq.com
*通讯作者:宋雪琳(1988—),女,讲师,研究方向:淡水鱼类分类与进化。E-mail:18271428161@163.com
鱲属(Zacco)隶属于鲤形目Cyprinifomes、鲤科Cyprinidae、马口鱼亚科Opsariichthyinae,是亚洲特有鱼类之一,在中国、日本、朝鲜、越南等均有分布[1]。该属鱼类喜栖息在山涧溪流中,个体较小,但种群数量较大,具有一定的经济价值[2]。近年来,随着我国经济的快速发展,水生态环境受到了严重的破坏,野生的鱲属鱼类的资源量正在逐渐减少。笔者查阅了该属鱼类的相关文献,对其分类、形态、生态学特征、遗传多样性等方面的研究材料予以综述,以期为深入研究、合理利用与保护该类鱼类资源提供参考。
1 鱲属的分类
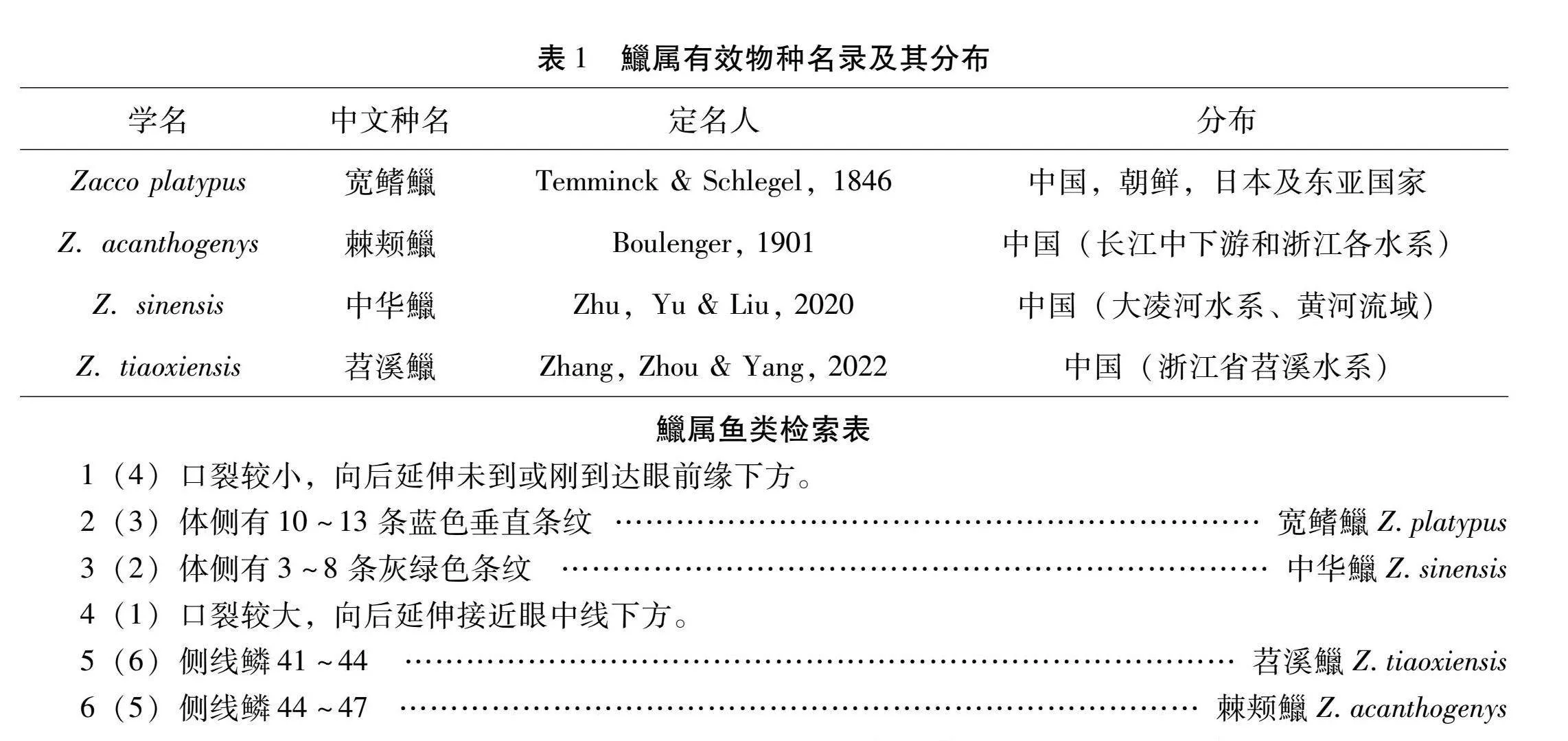
鱲属(Zacco)与其近缘属马口鱼属Opsariichthys、须鱲属Candidia和异鱲属Parazacco被分类学家归为马口鱼类[3],该类群以性成熟个体臀鳍条显著延长和明显的追星为主要特征,为我国溪河中的常见小型鱼类,其中以马口鱼属Opsariichthyinae和鱲属Zacco鱼类最为常见。Boulenger曾于1901年将采自浙江宁波的标本描述为马口鱼属新种棘颊马口鱼O. acanthogenys[4],该种随后被陈宜瑜归为宽鳍鱲的同物异名,并对马口鱼类的分类进行了重新整理,将Oshima[5]记录于台湾的纵纹鱲Z. temmincki描述为新种台湾鱲Z. taiwanensis,认为鱲属有宽鳍鱲Z. platypus、成都鱲Z. chengtui、纵纹鱲Z. temmincki、台湾鱲Z. taiwanensis和粗首鱲Z. pachycephalus 5个有效种[3]。
2008年,Chen等[6]基于线粒体D-loop序列的分析结果将纵纹鱲归入了新属东瀛鲤属Nipponocypris,随后又整合形态学特征和分子系统学的研究结果重新定义了鱲属鱼类,认为鱲属仅包含模式种宽鳍鱲,且该物种只分布于日本本岛,东亚大陆各地区记录的宽鳍鱲为同名异物,推测中国水系中可能有2~3个宽鳍鱲的近缘种。同时认为台湾鱲和粗首鱲为同物异名,且将其归入马口鱼属,有效种名为Opsariichthys pachycephalus[7]。Huang等[8]基于线粒体基因组全序列对鲤科鱼类的分类进行了重新整理,支持须鱲属Candidia和东瀛鲤属Nipponocypris的分类地位,并将成都鱲由鱲属归入马口鱼属。2015年,殷维[9]联合线粒体基因和核基因构建了马口鱼属的系统发育关系,在此基础上重新核验Boulenger对棘颊马口鱼的原始形态描述,认为该种为有效物种,并将其由马口鱼属归入鱲属,有效种名修订为棘颊鱲Z. acanthogenys (Boulenger,1901)。2019年,Wang等[10]以线粒体细胞色素b(Cyt b)基因为标记,在大量取样的基础上对马口鱼类进行了深入的系统发育研究,恢复了大鳞马口鱼(O. macrolepis)的分类地位,该种曾被认为是马口鱼的同物异名[3],认同Chen等[6]提出的宽鳍鱲只分布于日本,亚洲的样本为不同物种的观点。2020年,朱兰等[11]结合形态特征和分子序列,将采自辽宁大凌河和黄河水系的样本,即Wang等[10]研究结果中的一个谱系描述为新种中华鱲(Z. sinensis)。2022年,张琰等[12]基于形态和Cyt b基因全序列的系统发育关系分析,将采自于浙江苕溪水溪的鱲属标本描述为鱲属鱼类新种-苕溪鱲(Zacco tiaoxiensis sp.nov.)。因此,根据最新的分类系统,鱲属目前包括宽鳍鱲、棘颊鱲、中华鱲和苕溪鱲4个有效物种(表1)。本文根据相关文献资料编制了鱲属的检索表,并依据张琰等[12]基于Cyt b序列构建的马口鱼类系统发育树,归纳整理了马口鱼类及鱲属种间关系(图1)。
2 形态学研究
鱲属鱼类为溪流性小型鱼类,主要特征为:体长而侧扁,口端位,口裂向下倾斜,可达眼下方。上颌骨上有一缺口,下颌前端有一凸起与其相吻合。无口角须。侧线完全,在胸鳍上方显著下弯,至尾柄后回升到体侧中部。背鳍分枝鳍条7根,起点与腹鳍起点相对。臀鳍分枝鳍条9~10根,最长分枝鳍条向后延伸可达尾鳍基部[1]。根据Huang等[8]提出的分类系统,鱲属和马口鱼属主要的区别在于体侧条纹。鱲属鱼类体侧为较小的横条纹融合而成的不规则斑块,而马口鱼属鱼类体侧为独立而清晰的横条纹。
鱲属目前包括的4个有效种宽鳍鱲、棘颊鱲、中华鱲和苕溪鱲中,宽鳍鱲与中华鱲形态较为相似,棘颊鱲与苕溪鱲形态较为相似,但也都存在可区别的特征:宽鳍鱲与中华鱲两者在头长占比、背鳍占比、尾柄占比和鼻孔大小上存在差异[11];棘颊鱲与苕溪鱲可通过侧线鳞、侧线上鳞、侧线下鳞以及围尾柄鳞数目等可数性状区别开[12]。
3 生态学特征
目前有关鱲属鱼类生态学的报道较少,其中绝大部分都是以宽鳍鱲为研究对象的材料。
3.1 生活习性与食性
宽鳍鱲为喜栖息在山涧溪流中的群居小型鱼类[2]。宽鳍鱲广泛分布于长江和珠江流域的干流、支流以及一些附层水体中[10],尽管它的分布范围宽广,但主要是在山区河流,而且对水质的要求极高,一般生活在水流湍急、溶解氧含量高、pH呈中性、底质通常是沙石、水质洁净和有大量浮游植物生长的河里[13]。宽鳍鱲食性杂,常摄食甲壳类、藻类和有机碎屑等[14]。
3.2 年龄组成与生长
鱼类同一物种不同地理种群间的生活史特征往往存在很大差异[15-16],因此要根据不同地区来分析宽鳍鱲的年龄与生长特征。邢迎春等[13]研究表明北京地区的宽鳍鱲年龄组成较小,雄鱼最大3龄,雌鱼最大2龄;体长和体重相关关系式为:W=9×10-6L3.1947(R2=0.9784),其种群呈异速增长。项秀颖等[17]研究表明黄山区浦溪河的宽鳍鱲种群由Ⅰ、Ⅱ、Ⅲ共3种年龄组组成,雄鱼和雌鱼的最大年龄都为Ⅲ龄;雄鱼和雌鱼的体长和体重相关关系式不同:雄鱼的为W=6.26×10-6L3.1025(R2=0.9552),雌鱼的为W=4.88×10-6L3.1581(R2=0.9197),山区浦溪河宽鳍鱲两性间存在生长特性上的差异现象。严云志等[14]研究表明黄山地区徽水河的宽鳍鱲种群主要由1、2龄两个年龄组组成,雄、雌个体的最大寿命均为3龄;体长和体重相关关系式为:W=6×10-6L3.10(R2=0.9784)。曹安娜等[18]研究表明甬江溪口段宽鳍鱲种群由三个年龄组组成,其中2龄为优势年龄组;体长和体重相关关系式为:W=4×10-5L2.8035(R2=0.8552)。以上研究结果表明,由于分布地点不同,其生长环境有差异,造成了宽鳍鱲不同种群生长也有较大的差异。
3.3 繁殖特征
鱲属鱼类在繁殖生态上具有一定的相似性,个体通常1~2龄性成熟,繁殖期集中在4—7月,为春季产卵鱼类,一年只有一个繁殖季节。
由于生境不同,不同地区的宽鳍鱲种群在繁殖上表现出一定的差异性。就繁殖力而言,北京地区宽鳍鱲种群的绝对繁殖力的平均值为1715粒,相对繁殖力的平均值为532粒/g[13];黄山地区徽水河宽鳍鱲种群的绝对繁殖力为(758±362)粒,相对繁殖力为(77.38±22.15)粒/g[14];甬江溪口段宽鳍鱲种群的绝对繁殖力的平均值为(2580.39±2107.49)粒,相对繁殖力的平均值为(363.73±178.55)粒/g[18];青弋江宽鳍鱲种群的绝对繁殖力为(758±362)粒,相对繁殖力的平均值为(77.38±22.15)粒/g[19]。同时鱼类繁殖力与其生物学参数具有一定的相关性。孟子豪等[20]研究发现堵河宽鳍的绝对繁殖力与体长、体质量、空壳质量以及性腺质量均具有显著相关关系,与全长相关性不显著;曹安娜等[18]研究发现个体绝对繁殖力(F)与性腺重(WG)和年龄(A)有关,与WG正相关,与A负相关。
鱼类的繁殖关系到其种群能否维持稳定与发展,其特征是物种繁殖周期和外界环境条件共同作用的结果。环境条件的差异,如水温、光照和营养摄取等,会引起鱼类不同地理种群表现出不一样的性成熟时间、繁殖时间和繁殖力[21]。目前并没有研究专门针对宽鳍鱲不同种群繁殖力差异进行深入探讨,因此关于其不同种群间出现繁殖力差异较大的主要原因缺乏可靠的科学依据支持和具体的发生机制,这也是未来的一个重要研究方向。
4 遗传多样性
种群遗传多样性能够反映生物对环境变迁与自然选择的效应。种群遗传多样性越高,其演化潜力越大,适应生存环境能力越强,种群遗传多样性越低,承受的生存压力越大[22]。目前,我国已有一些宽鳍鱲自然群体遗传多样性的研究报道,一些学者采用线粒体DNA(mtDNA)序列对我国黑龙江、鸭绿江、海河、淮河、长江、珠江等东部和北部的大型河流中宽鳍鱲的系统地理学进行了初步研究,结果显示不同地区的宽鳍鱲发生了较大的遗传变异[23-25]。邢迎春等[26]利用mtDNA Cyt b基因研究了南方支系(长江、珠江)和北方支系(黑龙江、鸭绿江、海河、淮河)共计6条水系的宽鳍鱲系统地理学。梁晓旭等[27]基于mtDNA Cyt b基因对广东境内5条河流的宽鳍鱲种群遗传变异做了研究,认为不同地区的宽鳍鱲已产生了较大的遗传变异,随后又以mtDNA Cyt b基因全序列为分子标记,分析了广东境内9条河流的宽鳍鱲种群遗传变异和地理格局,认为广东地区宽鳍鱲群体出现了明显的分化。曹安娜[28]基于mtDNA COI和Cyt b序列对我国8个地区的宽鳍鱲群体进行了系统地理学研究,认为8个群体间的分布与地理格局之间无严格对应关系,但8个群体间因缺少基因交流,故而存在显著的遗传分化和群体间非随机交配。刘慧芬等[29]利用mtDNA COI基因序列作为分子标记,对河南境内四水系宽鳍鱲群体的群体分化和遗传多样性做了研究,认为宽鳍鱲的遗传变异多数存在于群体间。以上实验结果显示出了大部分宽鳍鱲种群具有较低的遗传多样性,但是宽鳍鱲种群的基因又存在明显的南北差异,这种差异既体现在控制鱼类外形特征的基因具有差异性,又体现在控制鱼类分子形态学特征上的基因具有差异性。
5 系统发育
基于线粒体细胞色素b(Cyt b)基因的系统发育研究显示,鱲属鱼类为单系群[11-12]。目前已被描述的4个有效物种中,棘颊鱲和苕溪鱲互为姐妹群[12],宽鳍鱲和中华鱲互为姐妹群[11]。经整理更新后,马口鱼类包含5个属:鱲属、马口鱼属、须鱲属、异鱲属和东瀛鲤属,鱲属与马口鱼属相比于其他3个属有更近的亲缘关系,二者互为姐妹类群。异鱲属鱼类处于系统发育树的基部,表明其较原始的地位。
6 鱼类资源现状
鱲属的4种鱼类中,以宽鳍鱲分布最广,广泛分布于中国绝大部分水域中;棘颊鱲分布于长江下游、钱塘江、甬江;中华鱲分布于黄河和大凌河水系[11];苕溪鱲目前仅分布在浙江苕溪水系[12]。虽然鱲属鱼类分布较广,但是其鱼类种群的遗传多样性处于较低水平,因自然因素和人为因素受到的生存压力也比较大。因此我们要继续加强对鱲属鱼类资源的保护,重点加强相关水域资源的养护,严格控制捕捞和环境污染,保护该属鱼类的优质资源。鱲属鱼类多在春季进行繁殖,当地渔业渔政管理部门应该在该时段严禁开展捕捞工作,严格打击禁渔期违法捕捞活动。此外,鱲属鱼类为急流性鱼类,喜栖息于河流支流的水流湍急处,拦河筑坝破坏了河流的连通性,改变了河流原来的水深、流速等水生生境,不利于鱲属及其他急流性鱼类的生存和繁殖[28]。Yan等[30]观察到低头坝建成后,其坝上蓄水区急流性鱼类种群规模显著减小,而缓流性和静水性鱼类显著增多。因此为有效保护鱲属鱼类的种质资源,还应加强对其栖息地的保护,降低水利工程建设、挖沙采矿等人类活动的威胁。
7 展望
鱲属鱼类有重要的经济价值和生态价值,属内物种如宽鳍鱲因繁殖季节体色艳丽,有很强的观赏价值,是颇受欢迎的原生观赏鱼类之一。鱲属鱼类目前已被描述的4个有效种中,除宽鳍鱲外,其余3个物种的有效地位已被广泛认可。其中中华鱲和苕溪鱲为近几年描述的新物种,分布区域较为狭窄,种群资源很容易受环境的影响。宽鳍鱲是本属据文献记录分布最为广泛的物种,也是分类地位最有争议的物种之一。目前多数学者认同该物种仅分布于模式产地日本,中国各水系分布的此前被鉴定为宽鳍鱲的类群应为宽鳍鱲的近缘种,这也说明鱲属鱼类的物种多样性被低估了,至少有2~3个隐存种有待描述。在今后的研究中,有望结合形态学、分子生物学和生态学特征对宽鳍鱲的分类地位进行深入分析,也为厘清马口鱼类这一类群的属级关系及属内种间关系提供依据。同时对本属鱼类进行深入研究,尤其是种群动态和繁殖生物学方面的研究,将为本属鱼类的合理利用和保护提供依据。
参考文献
[1] 乐佩琦.中国动物志-硬骨鱼纲-鲤形目(下卷), 鲤形目: 硬骨鱼纲 鲤形目[M].北京: 科学出版社, 2000: 40-46.
[2] 邢迎春,赵亚辉,唐文乔,等.中国宽鳍鱲Zacco platypus(Cypriniformes:Cyprinidae)的物种分化[C]//中国鱼类学会2008学术研讨会论文摘要汇编.南昌:中国鱼类学会,2008:60-61.
[3] 陈宜瑜.马口鱼类分类的重新整理[J].海洋与湖沼, 1982, 13(3): 293-299.
[4] Boulenger G A.Descriptions of new Freshwater Fishes discovered by Mr.F.W.Stynn at Ningpo, China[J].Proceedings of the Zoological Society of London, 1901, 70(2): 268-271.
[5] Oshima M.Contributions to the study of the fresh water fishes of the island of Formosa[J].Annals of the Carnegie Museum, 1919, 12: 169-328.
[6] Chen I S, Wu J H, Hsu C H.The taxonomy and phylogeny of Candidia (Teleostei: Cyprinidae)from Taiwan, with description of a new species and comments on a new genus[J].The Raffles Bulletin of Zoology, 2008, 19: 203-214.
[7] Chen I S, Wu J H, Hsu C H.The taxonomy and phylogeny of the cyprinid genus Opsariichthys Bleeker(Teleostei: Cyprinidae)from Taiwan, with description of a new species[J].Environmental Biology of Fishes, 2009, 86(1): 165-183.
[8] Huang S P, Wang F Y, Wang T Y.Molecular phylogeny of the Opsariichthys group(Teleostei: Cypriniformes)based on complete mitochondrial genomes[J].Zoological Studies, 2017, 56: e40.
[9] 殷维.马口鱼类的系统发育关系与生物地理研究[D].上海: 复旦大学, 2015.
[10] Wang X, Liu F, Yu D, et al.Mitochondrial divergence suggests unexpected high species diversity in the opsariichthine fishes(Teleostei: Cyprinidae)and the revalidation of Opsariichthys macrolepis[J].Ecology and Evolution, 2019, 9(5): 2664-2677.
[11] 朱兰, 俞丹, 刘焕章.中国北方鱲属鱼类一新种——中华鱲(鲤形目: 鲤科)[J].四川动物, 2020, 39(2): 168-176.
[12] 张琰, 周佳俊, 杨金权.中国南部鱲属鱼类一新种(鲤形目,鲤科)[J].上海海洋大学学报, 2023, 32(3): 544-552.
[13] 邢迎春, 赵亚辉, 张洁, 等.北京地区宽鳍鱲的生长及食性[J].动物学报, 2007, 53(6): 982-993.
[14] 严云志, 闫莉莉, 储玲, 等.徽水河宽鳍鱲的年龄、生长和繁殖[J].水生生物学报, 2012, 36(3): 474-481.
[15] Yan Y Z, Chen Y F.Changes in the life history of Abbottina rivularis in Lake Fuxian[J].Journal of Fish Biology, 2007, 70(3): 959-964.
[16] Yan Y Z, Chen Y F.Variations in reproductive strategies between one invasive population and two native populations of Pseudorasbora parva[J].Current Zoology, 2009, 55(1): 56-60.
[17] 项秀颖, 储玲, 周瑞龙, 等.黄山浦溪河宽鳍鱲的年龄和生长[J].淡水渔业, 2009, 39(6): 10-15.
[18] 曹安娜, 李强, 包薇红, 等.甬江溪口段宽鳍鱲的种群结构及繁殖力初步研究[J].水生态学杂志, 2019, 40(2): 94-101.
[19] 占姚军.青弋江两种亚科鱼类的生活史研究[D].芜湖: 安徽师范大学, 2011.
[20] 孟子豪, 李学梅, 王旭歌, 等.汉江支流堵河宽鳍鱲种群结构与繁殖力研究[J].四川动物, 2021, 40(3): 285-291.
[21] 殷名称.鱼类生态学[M].北京: 中国农业出版社, 1995.
[22] 韩增超.黑颈长尾雉圈养种群的遗传多样性分析[D].桂林: 广西师范大学, 2012.
[23] 梁晓旭, 庆宁, 杨柯林, 等.广东地区宽鳍鱲种群遗传变异和亲缘地理[J].水生生物学报, 2010, 34(4): 806-814.
[24] 闫锦锦.光唇鱼和宽鳍鱲线粒体全序列测定及分析[D].芜湖: 安徽师范大学, 2014.
[25] Xu Z, Primavera J H , Pena L D D L,et al.Genetic diversity of wild and cultured Black Tiger Shrimp(Penaeus monodon)in the Philippines using microsatellites[J].Aquaculture, 2001, 199(1-2): 13-40.
[26] 邢迎春.中国宽鳍鱲Zacco platypus(Cypriniformes: Cyprinidae)分子系统学、物种分化及其生物学特性的研究[D].武汉: 华中师范大学, 2007.
[27] 梁晓旭, 庆宁.广东地区不同水系宽鳍鱲种群遗传变异初探[C]// 中国鱼类学会2008学术研讨会论文摘要汇编.南昌:中国鱼类学会,2008: 43.
[28] 曹安娜.中国不同地区宽鳍鱲的系统地理学及甬江种群的生物学研究[D].广州: 中山大学, 2018.
[29] 刘慧芬, 张超, 王静, 等.河南境内四水系宽鳍鱲野生群体的遗传多样性[J].中国水产科学, 2018, 25(2): 269-277.
[30] Yan Y Z, Wang H, Zhu R, et al.Influences of low-head dams on the fish assemblages in the headwater streams of the Qingyi watershed, China[J].Environmental Biology of Fishes, 2013, 96(4): 495-506.
