顶坛花椒雄花花粉形态扫描电镜观测及生活力测定
2024-10-25罗辉曾钦朦卢中科侯娜娄丽王港
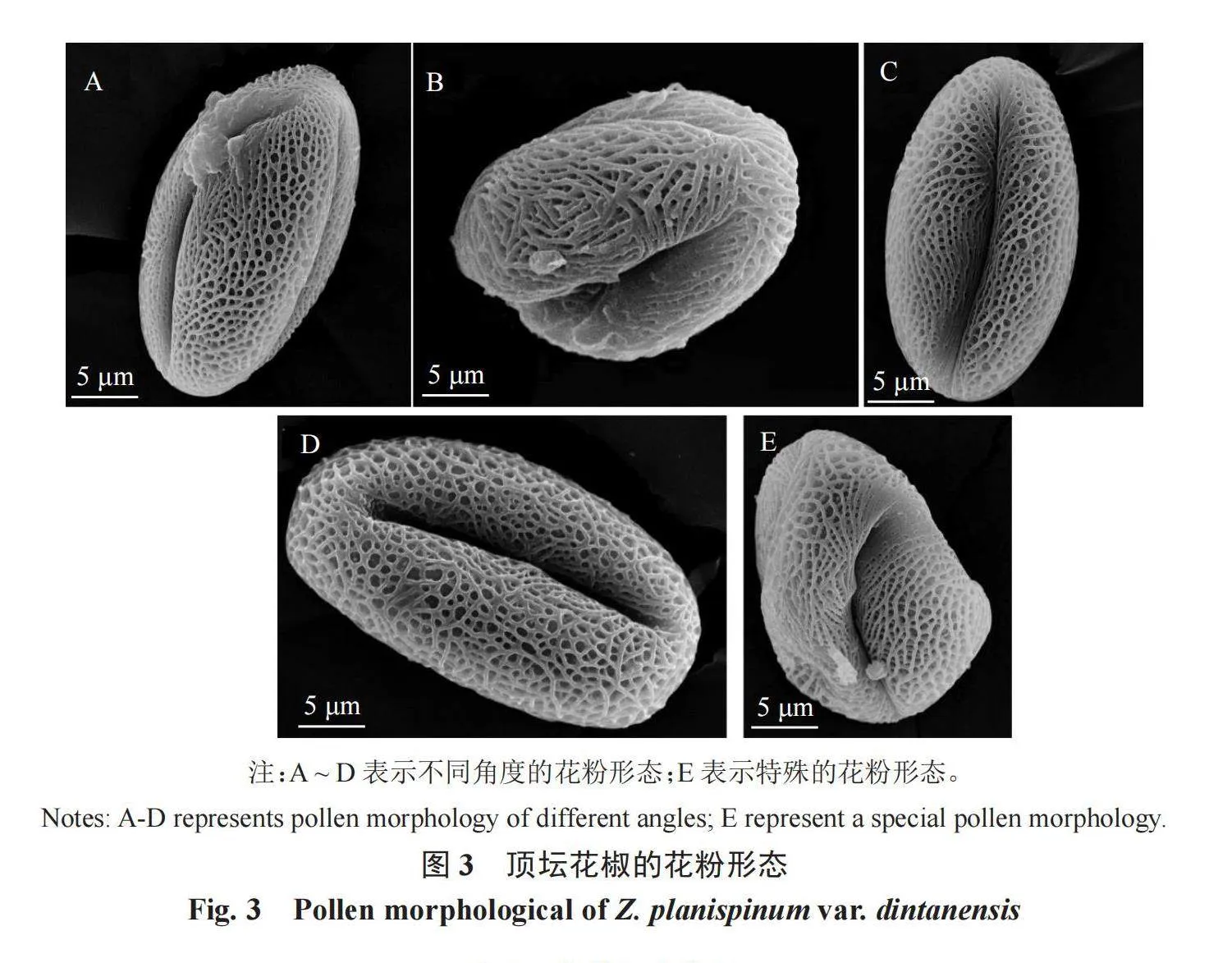



摘 要:以顶坛花椒花为试验材料,使用Eclipse E200生物显微镜和EM-30 Plus型扫描电镜对其花样品进行形态学观察和花粉活力分析,旨在为顶坛花椒杂交育种提供科学依据。结果表明,顶坛花椒出现雌雄同株的现象,花粉成熟后花药呈鲜黄色,花粉粒为长球形,表面具细网状纹饰。不同单株花粉粒直径存在显著差异,其中1号单株的花粉粒直径最大(522.32 μm),3号单株的花粉粒直径最小(279.50 μm)。花粉的极轴长、赤道轴长、花粉大小及网孔直径的平均长度分别为26.76、16.39、442.56和0.81 μm。花粉活力测试结果表明,1号和2号单株的花粉活力较高,亚历山大染色率分别为44.37%和48.57%,但植株长势较弱;5号单株的花粉活力较弱,亚历山大染色率仅有6.67%,但整个植株长势正常,无衰弱迹象。研究结果可为顶坛花椒繁殖策略的制定提供生物学依据。
关键词:顶坛花椒;形态构造;花粉粒大小;花粉活力
中图分类号:S573+.9 文献标志码:A 文章编号:1673-2871(2024)10-0118-06
收稿日期:2023-05-10;修回日期:2024-08-21
基金项目:贵州省科技厅中药现代化项目(黔科合ZY字〔2012〕3002号);贵州省林业厅项目(黔林科合2018〔01〕);贵州省科技厅成果转化项目(黔科合成果〔2019〕4233号);贵州省科技厅优秀青年科技人才项目(黔科合平台人才[20195643]);贵州省科技厅科技支撑项目(黔科合支撑〔2021〕222号,黔科合支撑〔2022〕一般118号)
作者简介:罗 辉,男,高级工程师,主要从事经济林栽培研究。E-mail:785486162@qq.com
通信作者:王 港,男,副研究员,主要从事林木遗传育种研究。E-mail:417328697@qq.com
Scanning electron microscopic observation and viability test of pollen grains of Zanthoxylum planispinum var. dintanensis
LUO Hui1, ZENG Qinmeng2, LU Zhongke2, HOU Na2, LOU Li2, WANG Gang2
(1. Guizhou Extension Station of Forestry Science and Technology, Guiyang 550001, Guizhou, China; 2. Guizhou Academy of Forestry Sciences, Guiyang 550005, Guizhou, China)
Abstract: The morphological observation and, pollen vitality analysis of the Zanthoxylum planispinum var. dintanensis flower were conducted using an Eclipse E200 biological microscope and an EM-30 Plus scanning electron microscope. This study aimed to provide a scientific basis for the hybrid breeding of Zanthoxylum bungeanum. The results showed that there was a phenomenon of monoecious growth in the Zanthoxylum planispinum var. dintanensis. Upon maturation, the anthers turned bright yellow, and the pollen grains were elongated spheres with fine mesh patterns on the surface. Significant variations were observed among different individual plants, specifically, the pollen grain diameter of No. 1 was the largest(522.32 μm), while that of No. 3 was the smallest(279.50 μm). The average length of the polar axis, equatorial axis, pollen size, and mesh diameter of the pollen were 26.76, 16.39, 442.56, and 0.81 μm, respectively. Pollen vitality tests revealed that No. 1 and No. 2 exhibited high vitality, with Alexander staining rate of 44.37% and 48.57%, respectively, although their associated plant growth was relatively weak. Conversely, the pollen vitality of Individual No. 5 was low, with an Alexander staining rate of only 6.67%, while the overall growth of the plant appeared normal with no signs of weakening. These research findings provide a biological basis for understanding the reproductive strategy of the Zanthoxylum planispinum var. dintanensis.
Key words: Zanthoxylum planispinum var. dintanensis; Morphological structure; Pollen size; Pollen vitality
顶坛花椒(Zanthoxylum planispinum var. dintanensis)是竹叶椒(Zanthoxylum planispinum)的一个变种[1],为芸香科花椒属植物,单性花,花序上所有小花均为雌花,无雄花,大多为纯雌株。顶坛花椒在贵州当地广为栽培,已成为重要的经济林木,对当地的生态环境修复发挥着重要作用[2]。近年来,许多学者对顶坛花椒开展了深入研究,主要集中在营养元素[3]、生理[4]、土壤养分[5]等领域,但对顶坛花椒花粉活力方面的研究较少。花粉是植物的特有器官,是植物种质保存、杂交育种及交换的重要材料[6]。不同植物的花粉经过演化和发展形成了独特的形态特征,即便是在同一种内的花粉,其生活力和形态也具有多样性[7]。花粉的形态特征主要受内环境的影响,通常由基因控制,具有较强的遗传稳定性及保守性[8],可为系统发育、识别和植物种类的鉴定提供科学依据,并在分类学及遗传学中广泛应用[9]。
无融合生殖是一种特殊的无性繁殖方式,在不经过雌雄配子融合的情况下产生种子,并能培育为成熟个体。这种生殖方式能够使植物的基因型保持不变,从而固定杂种优势,对作物育种具有极其重要的意义。无融合生殖在植物中的表现形式多样,包括营养的无融合生殖、无融合结子等类型,其遗传机制复杂,受多种途径调控[10]。花椒属植物普遍具有一个重要的生物学特性——无融合生殖,即雌性植株不需要经过受精,直接由珠心细胞发育成胚,进而繁殖后代[11]。刘映红等[12]研究表明,野花椒有正常的花粉,且花粉具有活力;叶萌等[13]对青花椒进行6年的观察,发现四川金阳地区的青花椒均为纯雌株,无雄花,而四川峨眉山市、洪雅县、雅安市雨城区有极少的藤椒存在雄花;李立新等[14]对花椒进行遗传多样性分析,发现由于花椒是无融合生殖方式,因此种群内变异较小。Fei等[15]通过细胞学观察、分子标记以及倍性鉴定等手段,进一步证明了花椒具有无融合生殖特性,并筛选出参与无融合胚胎发生的候选基因。此外,Wang等[11]对花椒无融合生殖的分子机制提出了新的见解,并基于基因组、转录组和代谢组技术对花椒属植物的两种生物碱的生物合成、无融合生殖及其抗旱特性进行了深入研究。
雌雄同株是指一个植物体上既有雌花又有雄花的现象,雌雄同株的植物可以自花授粉,也可以异花授粉,这种特性在植物繁殖策略中具有一定的优势[16]。笔者在进行顶坛花椒种质资源调查和品种选育的过程中,连续6年进行观察,发现顶坛花椒大多为纯雌株,部分顶坛花椒园中存在5%~10%着生雄花的植株,且出现雌雄同株的现象,同时花粉具有活力,这与上述学者的研究结果有所不同。鉴于此,笔者连续2年在顶坛花椒花期采集样品,采用生物学显微镜、扫描电镜及TTC染色法对具有代表性的顶坛花椒单株雄花形态构造、花粉形态指标及花粉活力等进行研究,以期为顶坛花椒的发育生物学理论及杂交育种提供科学依据。
1 材料与方法
1.1 试验地概况
试验地位于贵州省贞丰县北盘江镇,地处珠江水系西江上源北盘江畔,属喀斯特地貌地区,海拔550~1470 m,该地区属北亚热带高原季风湿润气候,气候温和,年均温16.4 ℃,全年无霜期335 d,雨量充沛,年均降水量1300 mm,雨热基本同季,年均日照时数1549 h,年均相对湿度81%,土壤以石灰土为主。
1.2 样品采集及保存
2021年3月至2022年3月,选择具有代表性的5株雌雄同株(编号1~5号)顶坛花椒,从树冠的不同方向采集正处于顶坛花椒盛花期的成熟花朵。将样品放置于恒温箱中干燥3 h,散粉后取下花粉,放置于铺有滤纸的培养皿中,于4 ℃冰箱保存备用。
试验地顶坛花椒不同单株见图1,其中1~4号(图1-A~D)单株的树龄为12年生,5号(图1-E)的树龄为2年生,在连续两年的观察中发现都有少数的雄花出现,且均为雌雄同株。1号顶坛花椒的植株长势较弱,表现出衰弱迹象,2号顶坛花椒一半有雄花,一半无雄花,同时发现开雄花的枝条长势比无雄花的枝条长势弱,3号和4号也出现同样的情况,但5号顶坛花椒幼株,开完雄花后植株长势正常。
1.3 研究方法
1.3.1 显微镜及扫描电镜观察 采用Eclipse E200生物学显微镜对花朵进行形态观察。将保存备用的样品用软毛刷轻微蘸取微量花粉,轻扫于电镜的固定液中,摇匀,在室温下固定2 h,再放置于4 ℃冰箱保存。固定好之后,使用0.1 mol·L-1的PB(磷酸)缓冲液清洗3次,每次15 min;依次经过不同浓度(φ,后同)的酒精(30%、50%、70%、80%、90%、95%、100%)进行脱水,再放置于干燥器皿内干燥,将干燥后的样品在ETD-900M磁控离子溅射仪中喷金30 s左右,置于EM-30 Plus超高分辨率台式扫描电镜下,选择有代表性的视野观察花粉的形态特征,随机选取10粒花粉进行测量。
1.3.2 花粉活力检测 采用亚历山大染色法进行花粉活力检测[17]。使用毛笔将花粉粒轻轻刷到载玻片上,并用滴管滴加亚历山大染料溶液染色5~10 h,然后在显微镜下观察、统计,原生质呈现红色且花粉壁呈现绿色的花粉具有活力,均呈现绿色的则为无活力或发育不良花粉。观察2~3张片子,每片取5个视野,统计花粉活力。花粉活力/%=红色花粉粒数/花粉粒总数×100。
1.4 数据分析
试验数据采用Excel 2007进行统计分析,用AutoCAD 2022测量花粉的形态指标,包括极轴长(P)、赤道轴长(E)、极赤比(P/E)和网孔直径,P×E为花粉的大小;参照《孢粉学概论》[6],0.88<P/E<1.14为圆球形,1.14≤P/E<1.33为近长球形,1.33≤P/E<2.00为长球形,P/E≥2.00为超长球形。利用Origin 2020b软件进行形态指标的差异显著性分析。
2 结果与分析
2.1 顶坛花椒花的形态构造及花粉粒大小
利用Eclipse E200生物显微镜对采集的样品进行观察,结果如图2所示。雌花的花小,花被片一轮,平均直径为3 mm,浅绿色(图2-A);子房上位有明显的花盘,花盘上着生2个离生心皮组成的雌蕊;子房为浅绿色,外侧向外凸起,且下部内侧相贴,呈花瓶状;花柱为闭合型,平均长620 μm,浅黄绿色,外倾且柱头二裂(图2-C)。雄花的大小与雌花相似,花被片一轮,8~10枚,绿色,有退化的雌蕊着生于花被片的内侧,6枚,花药背侧着生,为椭圆形,花粉成熟后的花药呈鲜黄色,花粉为黄色(图2-B、2-D~E)。
2.2 顶坛花椒花粉形态扫描电镜特征及分析
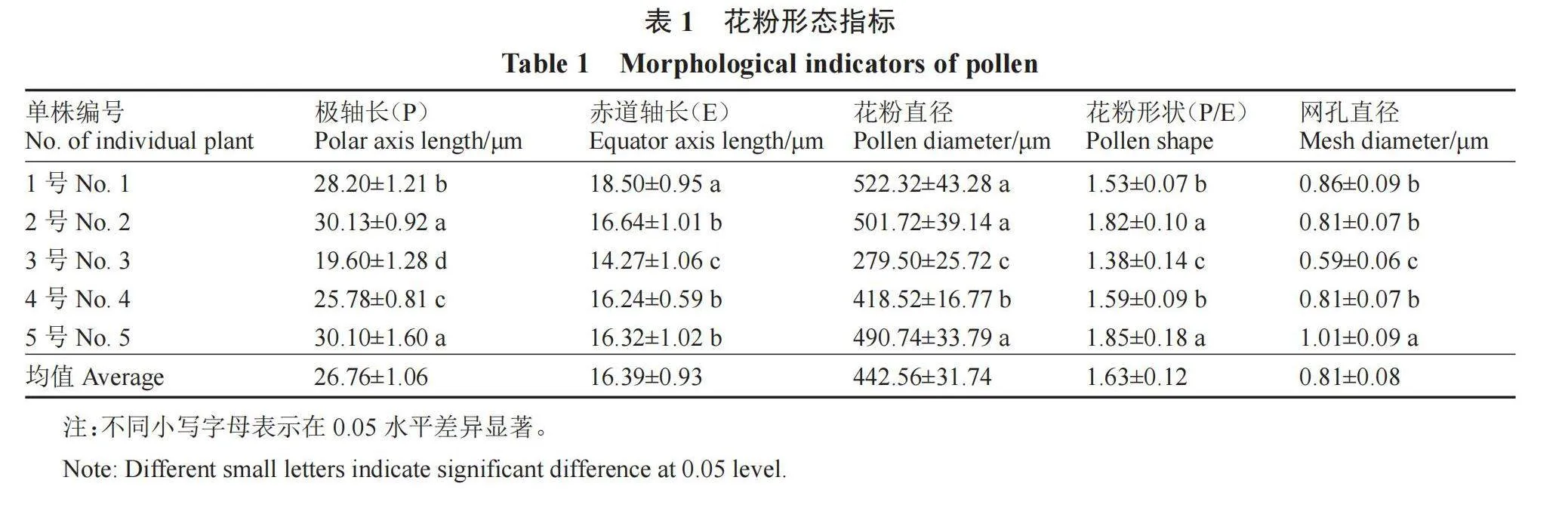
利用EM-30 Plus型扫描电镜对顶坛花椒不同单株的花粉形态进行观察,结果如图3和表1所示。花粉为长球形,偶有特殊的花粉形态,如图3-E,花粉的极面观为3裂的椭圆形,具孔沟,细长,勾线形,表面具有细网状纹饰,网孔呈不规则状。2号和5号单株的极轴长差异较小,显著高于其他单株,3号单株的极轴长最小,为19.60 μm;1号单株的赤道轴长为18.50 μm,显著高于其他单株。3号单株的花粉直径为279.50 μm,显著小于其他单株;1号单株的花粉直径最大,为522.32 μm。顶坛花椒5个单株花粉形状的P/E 值存在显著差异,但1.33<P/E<2.00,均为长球形。5号单株的网孔直径最大,为1.01 μm,显著大于其他单株;3号单株的网孔直径最小,为0.59 μm,显著小于其他单株。
2.3 顶坛花椒花粉活力检测
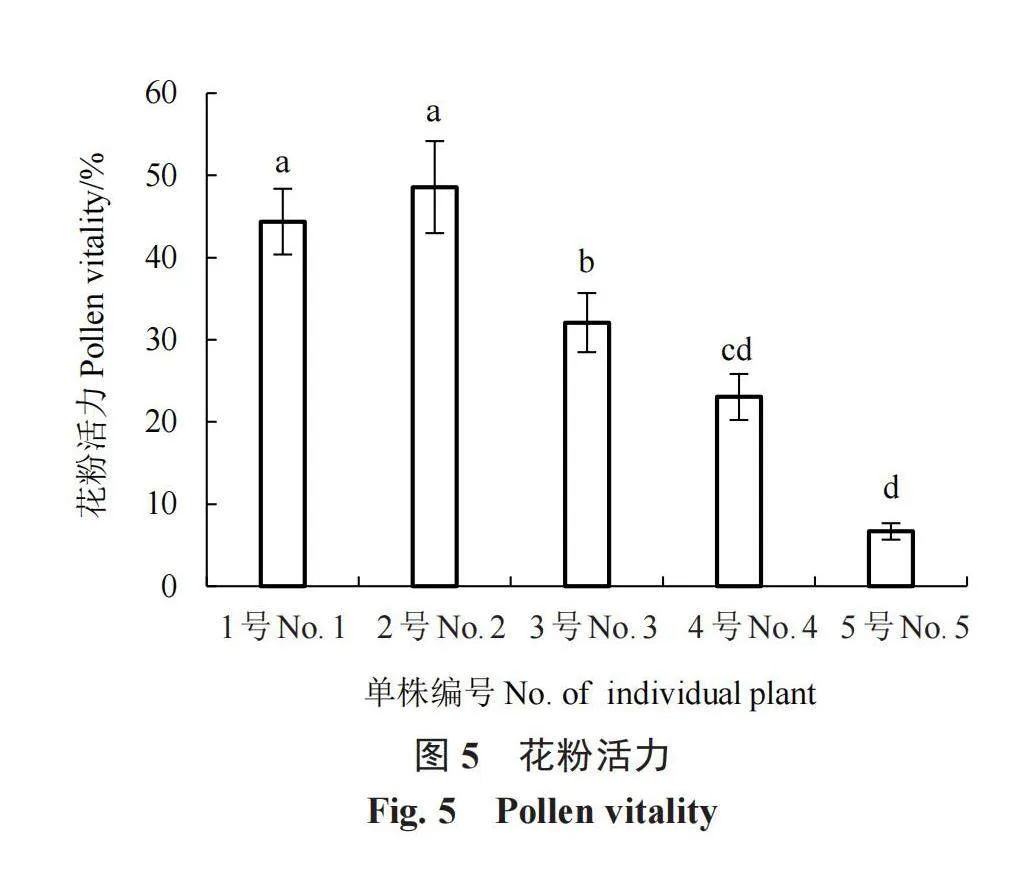
花粉活力是影响经济树种初始坐果率及产量的重要因素,顶坛花椒5个单株的花粉活力差异显著。从图4和图5可以看出,1号和2号顶坛花椒花粉活力经亚历山大溶液染色后呈红色的较多,分别为44.37%和48.57%,显著高于其他单株;5号顶坛花椒花粉亚历山大溶液染色后呈绿色的较多,花粉活力仅为6.67%。以上结果说明1号和2号顶坛花椒花粉活力较强,发育良好,而5号顶坛花椒多为发育不良或活力较弱的花粉。
3 讨论与结论
在《四川植物志》[18]、《贵州植物志》[19]和《浙江植物志》[20]等各地植物志中均写到竹叶花椒为单性花,在《福建植物志》[21]、《云南植物志》[22]和《中国植物志》[23]中描述了竹叶花椒的雌花和雄花。顶坛花椒是竹叶花椒的变种,均为无融合生殖;刘映红等[12]发现,北京栽培的花椒未找到雄株,也未见雄花,而野花椒为雌雄异株;而叶萌等[13]发现,栽培的竹叶花椒两个品种(藤椒和金阳青花椒)为雌花,具有无融合生殖特点,峨眉山市的藤椒园有雄花出现,但未做深入研究。笔者连续6年观察到试验地的顶坛花椒开出大量的“黄花”,且有雌雄同株的现象,这与刘映红等[12]的研究结果不一致。通过对顶坛花椒的实地及显微镜观察,并对花粉活力检测,发现1号和2号顶坛花椒单株的花粉活力最强,但其植株长势较弱,特别是1号顶坛花椒单株整株长势弱,有枯死迹象,而2号顶坛花椒单株左侧的枝条开雄花,右侧的枝条无雄花,发现无雄花的枝条长势比开雄花的枝条强,有可能是因为开雄花的枝条养分消耗较多,而无法正常供应整个枝条正常生长,导致植株长势衰弱,3号和4号顶坛花椒也表现出同样的现象。5号顶坛花椒整株开雄花后,整个植株长势正常,无衰弱迹象,出现这种现象可能是因为5号顶坛花椒单株是幼株,其生长能力旺盛,养分充足,且开出的雄花花粉活力差,因此开雄花对5号幼株的长势影响较小。
笔者对不同顶坛花椒单株的花粉形态指标进行测定,极轴长为19.60~30.13 μm,赤道轴长为14.27~18.50 μm,花粉直径为279.50~522.32 μm,这与刘炳伦[24]研究的花椒属的形态特征相似。开雄花的植株会出现植株生长衰弱的现象,但幼株并没出现这一现象。目前尚不清楚其发生原因,后续将进一步观察顶坛花椒产生雄花的现象是否与环境有关,对顶坛花椒雄花现象的深入研究对完善花椒的生殖发育理论和生产应用具有重要价值。
综上所述,笔者发现顶坛花椒出现雌雄同株现象,通过扫描电镜观察得出顶坛花椒花粉为长球形,偶有特殊形态,花粉的极面观为3裂的椭圆形,表面具有细网状纹饰,网孔呈不规则状。不同顶坛花椒单株的花粉活力存在差异,1号和2号单株的花粉活力较高,但植株长势较弱;5号单株的花粉活力较弱,但植株正常生长。
参考文献
[1] APPELHANS M S,REICHELT N,GROPPO M,et al.Phylogeny and biogeography of the pantropical genus Zanthoxylum and its closest relatives in the proto-Rutaceae group (Rutaceae)[J].Molecular Phylogenetics and Evolution,2018,126:31-44.
[2] 李红,喻阳华,龙健,等.顶坛花椒叶片功能性状对早衰的响应[J].生态学杂志,2021,40(6):1695-1704.
[3] 敖厚豫,李欣,余天华,等.基于麻味成分的顶坛花椒HPLC指纹图谱研究[J].中国调味品,2020,45(5):19-26.
[4] 李红,喻阳华.干热河谷石漠化区顶坛花椒叶片功能性状的海拔分异规律[J].广西植物,2020,40(6):782-791.
[5] 喻阳华,钟欣平,李红.黔中石漠化区不同海拔顶坛花椒人工林生态化学计量特征[J].生态学报,2019,39(15):5536-5545.
[6] 王开发,王宪曾.孢粉学概论[M].北京:北京大学出版社,1983.
[7] 尹佳蕾,赵惠恩.花粉生活力影响因素及花粉贮藏概述[J].中国农学通报,2005,21(4):110-113.
[8] SALMAKI Y,JAMZAD Z,ZARRE S,et al.Pollen morphology of Stachys (Lamiaceae) in iran and its systematic implication[J].Flora,2008,203(8):627-639.
[9] 武海霞,刘丽婷,廖柏勇,等.20种桉树及杂交种的花粉形态分析及分类学意义[J].中南林业科技大学学报,2012,32(3):29-36.
[10] 张燕,王春,王克剑.人工创制植物无融合生殖的研究进展[J].科学通报,2020,65(27):2999-3007.
[11] WANG M C,TONG S F,MA T,et al.Chromosome-level genome assembly of sichuan pepper provides insights into apomixis,drought tolerance,and alkaloid biosynthesis[J].Molecular Ecology Resources,2021,21(7):2533-2545.
[12] 刘映红,王伏雄,钱南芬.花椒和野花椒的无融合生殖[J].遗传学报,1987,14(2):107-113.
[13] 叶萌,杨德富,赵敏,等.青花椒的无融合生殖现象[C]//中国林学会.第三届中国林业学术大会论文集,2013.
[14] 李立新,司守霞,魏安智,等.基于花椒转录组序列SSR分子标记开发及花椒种质鉴定[J].华北农学学报,2017,32(5):69-77.
[15] FEI X T,SHI Q Q,QI Y C,et al.ZbAGL11,a class D MADS-box transcription factor of Zanthoxylum bungeanum,is involved in sporophytic apomixis[J].Horticulture Research,2021,8(1):23.
[16] 吴淋慧,董家琦,郑浩,等.利用猕猴桃雌雄花同株材料发掘性别决定相关基因[J].植物遗传资源学报,2024,25(7),1153-1162.
[17] ALEXANDER M P.A versatile stain for pollen,fungi,yeast and bacteria[J].Stain Technology,1980,55(1):13-18.
[18] 四川植物志编辑委员会.四川植物志:第九卷[M].成都:四川人民出版社,1981.
[19] 《贵州植物志》编辑委员会.贵州植物志:第二卷[M].贵阳:贵州人民出版社,1986.
[20] 韦直,何业祺.浙江植物志:第三卷[M].杭州:浙江科学技术出版社,1993.
[21] 福建省科学技术委员会《福建植物志》编写组.福建植物志:第二卷[M].福州:福建科学技术出版社,1985.
[22] 中国科学院昆明植物研究所.云南植物志:第六卷[M].北京:科学出版社,1995.
[23] 中国科学院中国植物志编辑委员会.中国植物志:第四十三卷第二分册[M].北京:科学出版社,1997.
[24] 刘炳仑.中国芸香科植物的花粉形态[J].植物研究,1987,7(3):11-37.
