长白落叶松自由授粉家系生长变异及优良家系早期选择
2024-10-09王佳兴闫平玉孙佰飞刘劲宏冯可乐张含国




摘要:【目的】通过对4个长白落叶松种子园的40个自由授粉家系在4个地点建立子代测定林,对其生长性状进行遗传变异和稳定性分析,初步选出长白落叶松优良家系。【方法】对来自鹤岗、林口、永吉、大孤家4个地点的长白落叶松家系6~8年生子代林树高、胸径进行单点方差分析、多年多点联合方差分析、遗传参数分析、稳定性分析及育种值估算,进行长白落叶松优良家系早期选择。【结果】单点方差分析表明,长白落叶松家系生长性状在家系间差异极显著;多年多点联合方差分析表明,长白落叶松家系树高性状在家系、地点、年份及互作间均差异极显著;遗传参数分析发现,不同地点不同年度树高家系遗传力均大于单株遗传力,各地点家系遗传力为0.611~0.852,4个地点联合家系遗传力为0.646,受到较强遗传控制。4个地点联合家系表型变异系数和遗传变异系数分别为41.36%和3.87%。20%入选率时8年生各地点树高性状遗传增益为23.35%~38.89%。以育种值估算结果为主,结合树高平均值和稳定性参数,选出CH309、CH349、HG5、BS349和HG13为高产稳产家系,平均育种值为0.528,平均树高为4.00 m,高出对照25.78%,平均稳定性参数为0.085,适宜在4个地点进行推广。【结论】长白落叶松家系生长性状在家系间、不同年度及不同地点间具有丰富的遗传变异,筛选出的优良家系适合在东北三省及立地条件相似的地区进行推广。
关键词:长白落叶松;生长性状;遗传变异;稳定性分析;优良家系;早期选择
中图分类号:S722"""""" 文献标志码:A开放科学(资源服务)标识码(OSID):
文章编号:1000-2006(2024)05-0081-09
Growth variation and superior families early selection of Larix olgensis free-pollinated families
WANG Jiaxing, YAN Pingyu, SUN Baifei, LIU Jinhong, FENG Kele, ZHANG Hanguo*
(State Key Laboratory of Tree Genetics and Breeding, Northeast Forestry University, Harbin" 150040, China)
Abstract:
【Objective】 We investigated genetic variation and stability in growth characteristics across 40 free-pollinated families from four Larix olgensis seed orchards located in different regions, aiming to identify superior families. 【Method】 We analyzed progenies from four L. olgensis locations namely Hegang, Linkou, Yongji" and Dagujia. The study involved single-point analysis of variance (ANOVA), multi-point multi-year ANOVA, genetic parameter analysis, stability analysis, and breeding value estimation on tree height and diameter at breast height (DBH) for trees aged 6-8 years to identify superior families. 【Result】 Single-point ANOVA results indicated significant differences in growth traits among families. Multi-point multi-year ANOVA revealed significant variations in the height of L. olgensis among families, locations, years, and their interactions. Genetic parameter analysis demonstrated that family heritability of height, ranging from 0.611 to 0.852 across sites, was greater than individual heritability, indicating strong genetic control. The phenotypic and genetic coefficients of variation at each site were 41.36% and 3.87%, respectively. Genetic gain in" eight year height ranged from 23.35% to 38.89% at a 20% selection rate. Breeding value estimation identified high-yielding and stable families (CH309, CH349, HG5, BS349, and HG13) with an average breeding value of 0.528, an average height of 4.00 m (25.78% higher than that of the control), and an average stability parameter of 0.085, making them suitable for promotion in four locations. 【Conclusion】 L. olgensis exhibits rich genetic diversity in growth traits among families across different years and locations. The identified superior families are well-suited for cultivation in the three northeastern provinces and similar environments.
Keywords:Larix olgensis; growth trait; genetic variation;stability analysis; superior family;early selection
林木生长性状作为林木最直观的表型性状,受遗传因素与环境因素共同作用[1-2]。我国落叶松遗传改良的起步时间始于20世纪60年代,取得了较多有价值的研究成果,得到了可信赖的信息及优良遗传材料。落叶松(Larix sp.)的生长性状是进行优良种源、家系或无性系选择的重要指标[3]。潘艳艳等[4]以67个日本落叶松(L. kaempferi)全同胞家系树高和地径为材料,得出日本落叶松全同胞家系苗期生长性状各变异来源差异均达极显著水平,树高和地径的表型变异系数分别为30.23%和21.51%,遗传变异系数分别为15.98%和16.99%;高扬等[5]在对8年生杂种落叶松(hybrid larch)8个家系的主要生长指标进行分析时发现,不同家系间的树高、胸径和当年高生长及材积的差异极显著 (Plt;0.01)。以上结果说明,根据表型选择得到的优树后代分化明显, 可以从中选择出生产力高的优良家系。
长白落叶松(L.olgensis)产于我国长白山区及老爷岭山区[6],具有生长快、分布广泛、适应性好、材质优良等特性,是东北地区防护林工程以及退耕还林、速生丰产林的重要造林树种。前期关于长白落叶松子代林遗传变异分析多数是利用单点数据,材料适应范围很难判断,开展多点遗传变异分析及稳定性分析有利于了解更广范围的遗传变异,并筛选出适应不同环境的优良材料,提高良种使用率。为此,本研究以黑龙江、吉林及辽宁省4个地点的长白落叶松自由授粉家系子代试验林为研究对象,对其进行遗传变异分析和稳定性分析,初步选择出长白落叶松优良家系,以期为深入开展落叶松遗传改良提供科学依据。
1 材料与方法
1.1 材料来源及试验设计
试验材料来自黑龙江省龙江错海(CH)、鹤岗(HG)、林口(LK)和宁安渤海(BS、NB)4个长白落叶松种子园的自由授粉家系,2013年播种育苗,2015年分别定植于黑龙江鹤岗林木良种繁育中心(130°26′56″E,47°32′59″N)、黑龙江林口县青山林场(130°16′32″E,45°16′32″N)、吉林永吉县西阳林场(126°17′46″E,43°35′56″N)和辽宁清原县大孤家林场(124°51′12″E,42°22′36″N)。鹤岗市林木良种繁育中心年均气温为3.8 ℃,年均降水量为651.5 mm;林口县青山林场年均气温为3.2 ℃,年均降水量为650 mm;永吉县西阳林场年均气温为4.0 ℃,年均降水量为700 mm;清原县大孤家林场年均气温为5.3 ℃,年均降水量为806.5 mm。
参试家系中来自鹤岗的65个、林口79个、永吉54个、大孤家82个,各地点设置当地对照各1个(鹤岗、永吉为当地长白落叶松种子,林口为长白落叶松种子园种子,大孤家为日本落叶松种子),4个地点共有相同家系40个。试验采用完全随机区组设计,4行32株小区,4次重复,株行距1.5 m×2.0 m。
1.2 数据调查与分析
于2018、2019、2020年秋季树木停止生长后调查子代测定林树高和胸径性状(分析仅采用保存率较高的3个重复,2018年林口生长性状未调查),根据拉依达准则[7]剔除数据异常值。
1)模型选择。单点方差分析模型[8]为
yjkl=μ1+Bj+Fk+(BF)jk+ejkl。(1)
式中:yjkl表示第j个区组、第k个家系内第l个单株的树高;μ1表示总体均值;Bj为区组的效应;Fk为家系的效应;(BF)jk为区组与家系间交互作用的效应;ejkl为随机误差效应。
长白落叶松年份、地点和家系间的3因素方差分析模型[9]为
Xijkl=μ2+τi+νj+(τν)ij+ωk+(τν)ik+(νω)jk+(τνω)ijk+ρjkl+εijkl。(2)
式中:Xijkl为第i个家系在第j地点、第k年份、第l区组的观测值;μ2为群体的平均值;τi为家系i的效应值;νj为地点j的效应;wk为年份的效应值;(τν)ij为家系×地点互作效应;(τν)ik为家系×年份的互作效应;(νω)jk为地点×年份的互作效应;(τνω)ijk为家系×地点×年份的互作效应;ρjkl为家系×地点×年份的互作效应;εijkl为随机误差。
2)遗传参数计算。包括家系遗传力、单株遗传力、表型变异系数、遗传变异系数和遗传增益。家系遗传力(h2F)和单株遗传力(h2S)[10]计算公式如下:
h2F=σ2F/(σ2Enb+σ2FBb+σ2F);(3)
h2S=4σ2F/(σ2E+σ2FB+σ2F)。(4)
式中:h2F为家系遗传力;h2S为单株遗传力;n为小区单株数;b为区组数;σ2F为家系方差分量;σ2E为机误;σ2FB为家系与区组互作的方差分量。
遗传变异系数(CCV,G)和表型变异系数(CCV,P)[11]公式为
CCV,G=σ2gX×100%;(5)
CCV,P=σ2PX×100%。(6)
式中:σ2g为遗传方差分量,σ2P为表型方差,X为性状的均值。
入选家系的遗传增益(ΔG)[12] 为
ΔG=(i×δ×h2F)/×100%。(7)
式中:i为选择强度(大小由入选率和群体大小决定,可通过正态分布表查得);δ为总体标准差;h2F为家系遗传力,为性状的均值。
3)AMMI(加性主效应乘积交互作用)模型是目前分析品种区域试验数据非常有效的模型[13-14]。本研究AMMI稳定性分析模型为
Ykij=μ+αk+βi+∑nk=1λrψkrσir+ρki+εkij。(8)
式中:Ykij为基因型k在环境i中第j次重复的观测值;μ是总体均值;αk是第k个基因型与总体均值的离差(即基因型主效应);βi是第i个环境与总平均的离差(即环境主效应);λr是第r个交互效应主成分轴(IPCA)的特征值或奇异值,代表交互作用平方和中由该轴解释的部分;ψkr是第r轴的基因型特征向量值;σir是第r轴的环境特征向量值;n为保留在模型中的主成分因子轴的总个数;ρki为提取n个IPCA轴后留下的残差(交互剩余项);εkij为试验误差。
家系与地点的相对稳定性参数(Dg)[13]为
Dg=∑nk=1ωnγ2kn。(9)
式中:n为提取的IPCA个数;ωn为权重系数,表示每个IPCA所解释的平方和占全部IPCA解释的平方和的比例;γ2kn为第k个基因型在第n个IPCA上的得分。Dg值越小,稳定性越高。
采用最佳线性无偏估计(BLUP)方法进行育种值估算。家系育种值(VB)[2]估算公式为
VB=Xβ+Zμ0+ε。
式中:β为区组效应,μ0为家系遗传效应(简称家系效应);ε为随机误差效应;X和Z分别为区组效应β和家系效应μ0的关联矩阵。
利用Excel、SPSS 22.0、DPS 14.10和R 4.1.2进行数据处理及分析。
2 结果与分析
2.1 长白落叶松家系生长性状差异分析
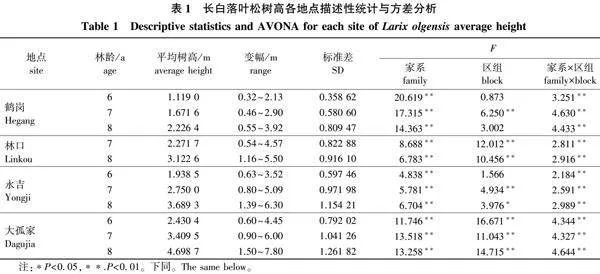
单点方差分析结果(表1)显示,树高性状在同一地点不同年份中,家系间以及家系与区组互作效应均显示差异极显著(Plt;0.01),说明家系间以及在小环境间生长差异明显。鹤岗地点8年生平均树高前5的家系高出当地对照117.21%,高出家系
平均值18.83%;林口地点8年生平均树高前5的家系高出当地对照18.81%,高出家系平均值21.47%;永吉地点8年生平均树高前5的家系高出当地对照33.24%,高出家系平均值26.02%;大孤家地点8年生平均树高前5家系低于当地对照8.5%,高于家系平均值16.81%。
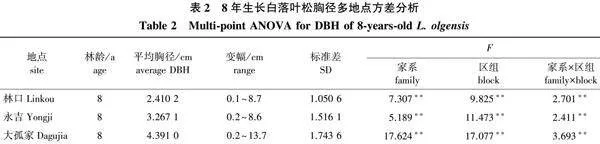
对8年生3个地点长白落叶松的胸径(鹤岗当年未调查)进行方差分析,结果如表2所示,3个地点的胸径性状在家系、家系与区组互作间均表现出差异极显著(Plt;0.01)。林口地点平均胸径前5的家系高出当地对照31.30%,高出家系平均值35.27%;永吉地点平均胸径前5的家系高出当地对照56.42%,高出家系平均值42.02%;大孤家地点平均胸径前5的家系低于当地对照22.88%,高于家系平均值22.55%。
以树高方差分析结果为基础,计算家系效应、区组效应以及家系×区组互作效应,并对各效应所占百分比进行分析可知:4个地点均主要受家系效应和家系区组互作效应影响。鹤岗地点6、7、8年生家系效应分别为65.98%、48.89%和47.86%,家系×区组互作效应分别为34.02%、50.57%和52.14%;林口地点2个年度的家系效应分别为50.01%和39.81%,家系×区组互作效应分别为46.48%和56.93%;永吉地点3个年度家系效应分别为52.66%、45.11%和44.10%,家系×区组互作效应分别为47.34%、53.15%和54.60%;大孤家地点3个年度家系效应分别为30.07%、33.29%和30.78%,家系×区组互作效应分别为66.28%、64.98%和67.11%。从整体上看,除大孤家年度效应差别不大外,其他3个地点均呈现随年龄增加家系效应逐渐降低、家系互作效应逐渐升高的规律。
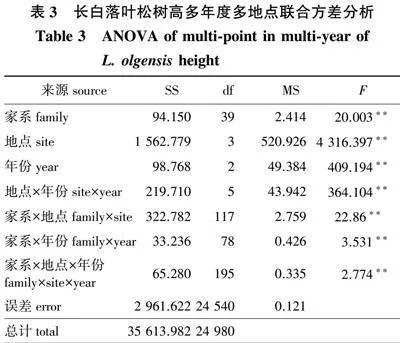
在4个地点误差均方同质的前提下,对4个地点共有的40个家系进行多年度多地点联合方差分析,结果(表3)发现,家系、地点、年份、地点×年份、地点×家系、年份×家系和地点×年份×家系互作间均表现出极显著差异(Plt;0.01),说明不同家系在同一地点、同一年份内生长差异明显;同一个家系在不同地点的生长和不同年份表现也各不一致,家系与地点、年份间存在较为明显的互作效应。
2.2 长白落叶松家系树高遗传参数分析
对4个地点3个年度长白落叶松树高性状的家系遗传力和单株遗传力进行估算,结果(图1a)发现:家系遗传力在各地点各年度数值均大于单株遗传力,且数值较高,说明不同地点不同年度树高性状均受到较强的遗传控制;鹤岗地点6~8年生家系遗传力随着林龄的增长略有下降,单株遗传力随着林龄的增长略有上升;林口地点7~8年生家系遗传力随着林龄的增长有所下降,单株遗传力随着林龄的增长也逐渐下降;永吉地点6~8年生家系遗传力随着林龄的增长略有下降,单株遗传力随着林龄的增长逐渐上升;大孤家地点家系遗传力随着林龄的增长呈现先上升后下降的趋势,但变化不大,单株遗传力随着林龄的增长逐渐上升。总体来看,家系遗传力随林龄增加略有降低,单株遗传力除林口地点之外,其他地点随林龄增加逐渐升高。
对4个地点长白落叶松树高3年的表型变异和遗传变异进行分析,结果(图1b)发现,表型变异系数在不同地点不同年度均大于遗传变异系数。鹤岗地点6~8年生表型变异系数随着林龄的增长逐渐增加,遗传变异系数随着林龄的增长呈现先下降后上升的趋势;林口地点7~8 年生表型变异系数随着林龄的增长逐渐下降,遗传变异系数随着林龄的增长逐渐下降;永吉地点6~8年生表型变异系数随着林龄的增长呈现先上升后下降的趋势,遗传变异系数随着林龄的增长逐渐下降;大孤家地点6~8年生表型变异系数随着林龄的增长逐渐下降,遗传变异系数随着林龄的增长逐渐下降。
3个地点长白落叶松8年生胸径性状的遗传力和变异系数分析结果表明,家系遗传力均大于单株遗传力,表型变异系数均大于遗传变异系数。
对8年生4地点40个共有家系进行联合遗传参数计算发现,与单点类似,多点家系遗传力大于单株遗传力,分别为0.646和0.224;表型变异系数大于遗传变异系数,分别为41.36%和3.87%。
试验材料的遗传差异随着年龄的增长更能体现出来,所以对8年生各地点树高性状进行遗传增益估算。入选率为20%时,选择强度为1.4,遗传增益为23.35%~38.89%。
2.3 长白落叶松家系多点稳定性分析
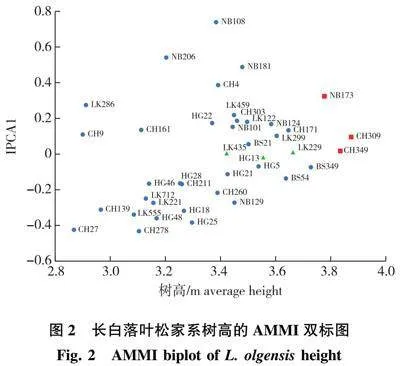
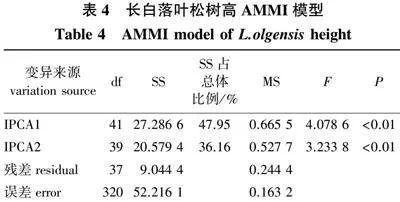
因树高性状的家系间和地点间以及家系与地点互作间达到差异极显著(Plt;0.01),对4个地点8年生长白落叶松树高性状进行AMMI模型回归分析,结果(表4)发现,第1主成分分量(IPCA1)能够解释交互作用的47.95%,前2项主成分分量能够解释交互作用的84.11%,且均达到差异极显著(Plt;0.01),说明应用AMMI模型来解释家系和地点的互作效应可行,用来分析家系树高稳定性的可靠程度较高。
为了进一步分析家系树高性状的稳定性,以家系地点树高性状平均值为X轴,第1主成分分量(IPCA1值)为Y轴做出AMMI 双标图(图2)。图中X轴数值越大,表示树高越大,Y轴绝对值越大,表示稳定性越差。由图2可以看出,在X轴方向上,CH309、CH349、NB173家系绝对值较大,生长相对性较好;CH27、CH9、LK286、CH139家系绝对值较小,生长相对性较差。在Y轴方向上,NB108、NB206、NB181、CH278等家系的IPCA1绝对值较大,说明这些家系与环境有着很强的交互作用,稳定性较差;而HG13、LK229、LK435等家系的IPCA1绝对值较小,说明环境对其影响较小,稳定性较好。进一步分析发现,地点间的分散程度要大于家系间的分散程度,说明环境影响也较大,同一家系在不同地点的生长表现差异较大。
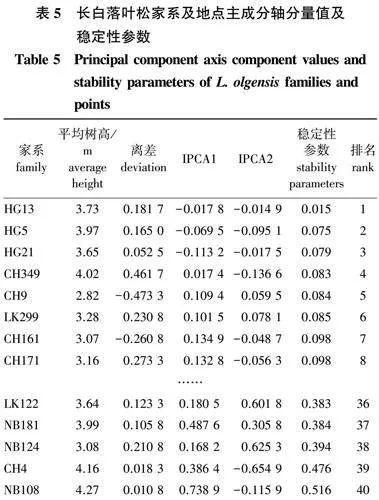
为了更好地比较各个家系的稳定性,计算各家系的稳定性参数,结果如表5所示。由表5可知,稳定性参数(Dg)越低,家系稳定性越好。稳定性小于0.1的有8个家系,稳定性最好的5个家系为HG13、HG5、HG21、CH349和CH9,平均稳定性参数为0.067。稳定性大于0.3的家系有10个,最差的5个家系为LK122、NB181、NB124、CH4和NB108,平均稳定性参数为0.431。
2.4 长白落叶松家系育种值估算及优良家系选择
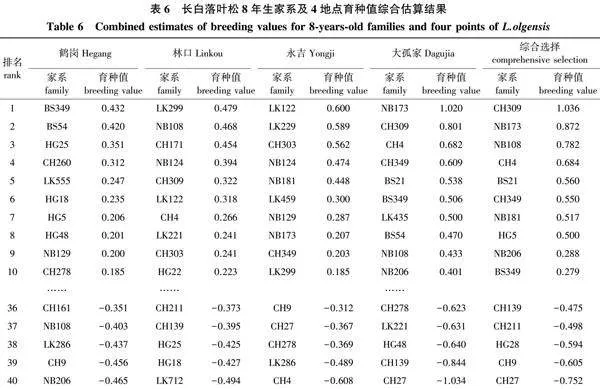
对4个地点共有的40个长白落叶松家系进行育种值估算,并对4个地点8年生家系育种值进行排名,结果见表6。鹤岗地点8年生育种值前5家系平均育种值为0.353,高出对照169.30%;林口地点8年生育种值前5家系平均育种值为0.424,高出对照1 033.42%;永吉地点前5家系平均育种值为0.535,高出对照405.25%;大孤家地点前5家系平均育种值为0.731。总体看,各年度各地点家系育种值排名虽有小幅波动,但基本保持稳定状态。
对8年生4个地点的家系育种值进行综合估算,育种值大于0的家系共有18个,其中排名前5的家系分别为CH309、NB173、NB108、CH4和BS21,平均育种值为0.787,排名后5的家系为CH139、CH211、HG28、CH9和CH27,平均育种值为-0.585。
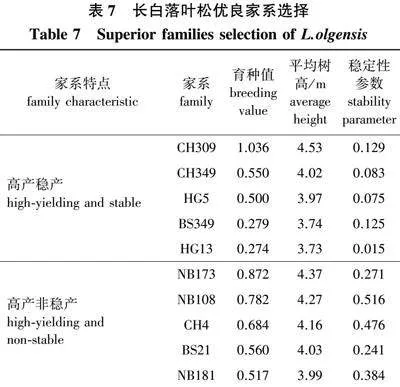
以育种值估算结果为主,结合树高平均值和稳定性参数,在育种值大于0的18个家系中选择优良家系,结果见表7。其中高产稳产的家系有CH309、CH349、HG5、BS349和HG13,平均育种值为0.528,平均树高为4.00 m,高出对照25.78%,平均稳定性参数为0.085,适宜在4个地点进行推广。高产非稳产的家系有NB173、NB108、CH4、BS21和NB181,平均育种值为0.683,平均树高为4.16 m,高出对照30.82%,平均稳定性参数为0.377 8。其中BS21家系适宜在黑龙江小兴安岭地区进行推广,NB108、CH4、BS21和NB173家系适宜在黑龙江省张广才岭和老爷岭地区进行推广。NB181和NB173适宜在吉林哈达岭地区进行推广。NB173、CH4、BS21和NB181家系适宜在辽宁长白山山脉地区进行推广。
3 讨 论
林木的遗传和变异研究是开展遗传改良的前提,掌握遗传变异规律是制定育种策略的基础[15]。林木生长性状变异受一定程度的遗传控制,有效的遗传变异决定了遗传改良的潜力,选择优良的家系和单株用于营建种子园生产良种并加以推广应用,是良种选育和生产相结合的首选方式[16]。研究表明,具有相同遗传结构的树种在个体、家系和地点间均存在较大的遗传变异,不同的环境和栽培措施下生长性状具有差异性[17]。本研究发现,长白落叶松家系生长性状在家系、地点、年龄及其互作间均存在极显著的差异,说明长白落叶松家系生长性状存在丰富的变异基础,这与苑海静等[1]对麻栎生长性状的研究结果一致。林木间的生长差异是在遗传和环境的共同作用下形成的,长白落叶松家系树高性状受家系效应和家系区组互作效应的影响,并且除大孤家地点外,其余3个地点随着树龄的增长,家系效应下降,家系与区组互作效应上升,说明随着树龄增加,环境影响也在逐渐上升。因此,在生产上应针对立地环境选用适生的品种。此外,由于我国长白落叶松种子园仍然存在产量不足和遗传增益较低等问题,所以可进行后向选择,通过选择出的优良家系来指导种子园管理以及优树的选择。刘宇等[18]利用家系育种值评估了12年生白桦子代测定林,指导了种子园亲本的伐劣留优,提高了种子园良种品质;金国庆等[19]通过对马尾松一代和二代自由授粉家系进行育种值估算及生长评价,解决了种子园建园亲本的再选择和留优去劣疏伐现有种子园。
精确估算遗传参数对于预期遗传增益、早期选择和科学制定育种策略具有指导意义[20-22]。林木性状遗传力估算值不仅因性状而有差异,还与参试材料的数量及试验材料对环境的适应程度有关,林龄、所处生境和试验设计等条件的不同也会导致遗传力的差异。较强的遗传控制有利于获得最大化的遗传增益效果[23-24]。Diao等[25]发现日本落叶松家系在4~18年间树高、胸径的家系和单株遗传力随年龄增长呈一定的动态变化,家系遗传力大于单株遗传力。李安琪等[26]对30个黑木相思半同胞家系进行研究时发现,生长性状的家系遗传力均大于单株遗传力,受较高遗传控制。本研究发现,各地点各年度家系遗传力范围为0.611~0.852,说明树高性状受到较强的遗传控制;单株遗传力范围为0.275~0.597。各个年度家系遗传力均大于单株遗传力,说明选择优良家系可以获得更多的遗传增益。单株遗传力小于家系遗传力可能是因为受地区环境影响,家系内单株变化较大的缘故。但若能将家系选择和单株选择相结合,可能会取得更好的改良效果。进一步研究发现,家系遗传力随着林龄的增加,总体上呈现下降的趋势,这与唐良民等[27]对马尾松的研究结果相似,可能是因为随着林龄的增加,环境作用的累积效应加强,导致受到的遗传控制减弱。此时,受基因与环境效应作用,家系整体表现逐渐稳定,不同年度的遗传力变化幅度逐渐减小。对长白落叶松各地点各年份树高性状的遗传变异系数和表型变异系数进行计算时发现,各地点各年度表型变异系数均大于遗传变异系数,且差距较大,这与潘艳艳等[4]对日本落叶松家系的研究结果一致。表型是在遗传和环境的共同作用下形成的,说明在生长初期,环境对于家系变异的影响较大。另外,各地点各年度树高性状的表型变异系数和遗传变异系数均处于波动状态,表型性状分化较大,说明在生长初期,树高性状受遗传效应引起变异的变化趋势是不同的。
开展多点试验对参试家系进行稳定性与适应性分析一直备受育种工作者的广泛关注。过去有关林木稳定性分析多采用回归分析模型来评价,但该方法对环境效应与遗传效应不是独立估计,并且基因型与环境的互作效应也并非简单线性叠加,所以回归分析模型不是一种理想的互作效应分析方法[28]。在Eberhart和Russell模型中,实际测量的数据,家系的离回归均方与0有着显著差异,模型评价不可靠,这与张磊[29]对杂种落叶松家系进行稳定性分析时使用Eberhart和Russell模型所得结论一致。Shukla模型中家系的F值大部分是不显著的,可通过F值评价其稳定性,但结果方差值较大,评价精度不高,这与李火根等[30]对美洲黑杨使用Shukla模型分析结果一致。采用AMMI模型、George模型所得到的稳定性基本一致,并且与各地家系的真实表现相一致。所以在评价长白落叶松时应以AMMI模型为主[31]。刘宇等[32]利用AMMI模型在对白桦进行生长稳定性分析时发现,仅第1主成分分量就能够解释81.48%的交互作用,且达到了极显著水平(Plt;0.01)。李红盛等[33]利用AMMI模型对山苍子家系进行稳定性分析时发现,两项主成分分量能够解释交互作用的100%。本研究发现,2项主成分分量合计能够解释交互作用的84.11%,说明用AMMI模型来分析家系树高稳定性较可靠。对家系稳定性参数进行计算时发现,部分生长较好的家系稳定性却很差,如CH4和NB108家系的树高排名分别在第4和第3位,稳定性分析却只排在第39和40位,说明仅凭稳定性参数去选择优良家系是片面的,还应当结合生长性状表型值去选择。
育种值是性状表型值中遗传效应的加性效应部分,反映了遗传效应的大小,对林木育种值的精确预测是提高选育效率的关键[34]。所以本研究对4地点8年生长白落叶松家系树高育种值进行综合分析,并结合树高平均值以及稳定性参数,分别选出高产稳定型家系和高产非稳定型家系。由于育种目标的不同,对于后续长白落叶松家系的推广方向也应有所不同;对于高产稳定型家系,适宜在所有地区进行推广;对于高产非稳定型家系,应遵循适地适树原则。
总之,本研究中长白落叶松家系生长性状在家系间、不同年度间及不同地点间具有丰富的遗传变异,具有很大的选择空间。20%入选率时遗传增益在23.35%~38.89%之间变化。8年生4个地点40个共有家系的家系遗传力和单株遗传力分别为0.646和0.224,表型变异系数和遗传变异系数分别为41.36%和3.87%。选出5个高产且稳定的优良家系为CH309、CH349、HG5、BS349和HG13,家系树高平均值高出对照25.78%,可在东北三省及立地条件相似的地区推广。通过观察发现来自渤海种子园的自由授粉家系育种值排名比较靠前,种子园整体水平较高,对于推广混系良种的增产效果较好。
参考文献(reference):
[1]苑海静,成向荣,虞木奎,等.麻栎优树自由授粉家系生长性状3地点间动态变异及优良家系选择[J].林业科学研究,2022,35(2):9-18.YUAN H J,CHENG X R,YU M K,et al.Dynamic variation of growth traits in open-pollinated families of Quercus acutissima superior tree and selection of superior families among three sites[J].For Res,2022,35(2):9-18.DOI: 10.13275/j.cnki.lykxyj.2022.02.002.
[2]贾庆彬,刘庚,赵佳丽,等.红松半同胞家系生长性状变异分析与优良家系选择[J].南京林业大学学报(自然科学版),2022,46(4):109-116.JIA Q B,LIU G,ZHAO J L,et al.Variation analyses of growth traits in half-sib families of Korean pine and superior families selection[J].J Nanjing For Univ (Nat Sci Ed),2022,46(4):109-116.DOI: 10.12302/j.issn.1000-2006.202107040.
[3]徐贵友,张义飞,薛明旭,等.落叶松遗传改良研究进展[J].林业科技情报,2019,51(4):65-68.XU G Y,ZHANG Y F,XUE M X,et al.Research progress of genetic improvement of Larix spp.[J].For Sci Technol Inf,2019,51(4):65-68.DOI: 10.3969/j.issn.1009-3303.2019.04.026.
[4]潘艳艳,许贵友,董利虎,等.日本落叶松全同胞家系苗期生长性状遗传变异[J].南京林业大学学报(自然科学版),2019,43(2):14-22.PAN Y Y,XU G Y,DONG L H,et al.Genetic variations of seedling growth traits among full-sib families of Larix kaempferi[J].J Nanjing For Univ (Nat Sci Ed),2019,43(2):14-22.DOI: 10.3969/j.issn.1000-2006.201803038.
[5]高扬,王有菊,杨世桢,等.杂种落叶松优良家系的选择[J].中南林业科技大学学报,2013,33(10):57-60.GAO Y,WANG Y J,YANG S Z,et al.Superior family selection of hybrid larch[J].J Cent South Univ For Technol,2013,33(10):57-60.DOI: 10.14067/j.cnki.1673-923x.2013.10.025.
[6]中国科学院中国植物志编辑委员会.中国植物志:第七卷[M].北京:科学出版社,1978.
[7]张敏,袁辉.拉依达(PauTa)准则与异常值剔除[J].郑州工业大学学报,1997(1):84-88.ZHANG M,YUAN H.The PauTa criterion and rejecting the abnormal value[J].J Zhengzhou Univ Technol,1997(1):84-88.
[8]孙英豪,张含国,郝俊飞,等.3年生长白落叶松高生长遗传变异与多点稳定性[J].东北林业大学学报,2018,46(8):1-7.SUN Y H,ZHANG H G,HAO J F,et al.Genetic variation and stability of three-year growth height of Larix olgensis[J].J Northeast For Univ,2018,46(8):1-7.DOI: 10.13759/j.cnki.dlxb.2018.08.001.
[9]唐启义.DPS数据处理系统:专业统计及其他[M].3版.北京:科学出版社,2013.TANG Q Y.DPS data processing system[M].3rd ed.Beijing:Science Press,2013.
[10]蒋开彬,杜澄举,李赛楠,等.4年生火炬松半同胞家系生长和分枝性状遗传评估[J].北京林业大学学报,2020,42(9):1-10.JIANG K B,DU C J,LI S N,et al.Genetic evaluation on growth and branching traits of 4-year-old half-sib families of loblolly pine[J].J Beijing For Univ,2020,42(9):1-10.
[11]王云鹏,张蕊,周志春,等.10年生木荷生长和材性性状家系变异及选择[J].南京林业大学学报(自然科学版),2020,44(5):85-92.WANG Y P,ZHANG R,ZHOU Z C,et al.A variation and selection of growth and wood traits for 10-year-old Schima superba[J].J Nanjing For Univ (Nat Sci Ed),2020,44(5):85-92.DOI: 10.3969/j.issn.1000-2006.202003086.
[12]莫家兴,华慧,翁怀峰,等.柳杉全同胞家系生长和材性的遗传变异及优良家系选择[J].中南林业科技大学学报,2019,39(10):40-47.MO J X,HUA H,WENG H F,et al.Growth and wood property genetic variation analysis and superior family selection of Cryptomeria full-sib family[J].J Cent South Univ For Technol,2019,39(10):40-47.DOI: 10.14067/j.cnki.1673-923x.2019.10.007.
[13]解懿妮,刘青华,蔡燕灵,等.5年生马尾松生长性状3地点家系变异及评价[J].林业科学研究,2020,33(5):1-12.XIE Y N,LIU Q H,CAI Y L,et al.Family variation and evaluation of growth traits of 5-year-old Pinus massoniana in three sites[J].For Res,2020,33(5):1-12.DOI: 10.13275/j.cnki.lykxyj.2020.05.001.
[14]吴世雄,刘艳红,张利民,等.不同产地东北红豆杉幼苗迁地保护的生长稳定性分析[J].北京林业大学学报,2018,40(12):27-37.WU S X,LIU Y H,ZHANG L M,et al.Growth stability analysis of ex situ conservation of Taxus cuspidata seedlings from different sources[J].J Beijing For Univ,2018,40(12):27-37.DOI: 10.13332/j.1000-1522.20180331.
[15]MWASE W F,SAVILL P S,HEMERY G.Genetic parameter estimates for growth and form traits in common ash (Fraxinus excelsior) in a breeding seedling orchard at Little Wittenham in England[J].N For,2008,36(3):225-238.DOI: 10.1007/s11056-008-9095-6.
[16]WHITE T L,ADAMS W T,NEALE D B.Forest genetics[M].Wallingford,Oxfordshire,UK:CABI Pub,2007.
[17]ZHANG Z,JIN G Q,FENG Z P,et al.Joint influence of genetic origin and climate on the growth of Masson pine (Pinus massoniana Lamb.) in China[J].Sci Rep,2020,10(1):4653.DOI: 10.1038/s41598-020-61597-9.
[18]刘宇,徐焕文,张广波,等.白桦半同胞子代多点生长性状测定及优良家系选择[J].北京林业大学学报,2017,39(3):7-15.LIU Y,XU H W,ZHANG G B,et al.Multipoint growth trait test of half-sibling offspring and excellent family selection of Betula platyphylla[J].J Beijing For Univ,2017,39(3):7-15.DOI: 10.13332/j.1000-1522.20160154.
[19]金国庆,张振,余启新,等.马尾松2个世代种子园6年生家系生长的遗传变异与增益比较[J].林业科学,2019,55(7):57-67.JIN G Q,ZHANG Z,YU Q X,et al.Comparisons of genetic variation and gains of 6-year-old families from first-and second-generation seed orchards of Pinus massoniana[J].Sci Silvae Sin,2019,55(7):57-67.DOI: 10.11707/j.1001-7488.20190706.
[20]BALTUNIS B S,GAPARE W J,WU H X.Genetic parameters and genotype by environment interaction in Radiata pine for growth and wood quality traits in Australia[J].Silvae Genet,2010,59(1/2/3/4/5/6):113-124.DOI: 10.1515/sg-2010-0014.
[21]李光友,徐建民,李昌荣,等.杂交桉家系在桂北生长及优良性评价[J].中南林业科技大学学报,2021,41(2):8-15.LI G Y,XU J M,LI C R,et al.Study on the grouped and genetic analysis of Eucalyptus hybrids in northern Guangxi[J].J Cent South Univ For Technol,2021,41(2):8-15.DOI: 10.14067/j.cnki.1673-923x.2021.02.002.
[22]孙晓梅,张守攻,侯义梅,等.短轮伐期日本落叶松家系生长性状遗传参数的变化[J].林业科学,2004,40(6):68-74.SUN X M,ZHANG S G,HOU Y M,et al.Age trends of genetic parameters for growth traits in short rotation Larix kaempferi families[J].Sci Silvae Sin,2004,40(6):68-74.DOI: 10.3321/j.issn:1001-7488.2004.06.012.
[23]张谦,曾令海,蔡燕灵,等.樟树自由授粉家系生长与形质性状的遗传分析[J].中南林业科技大学学报,2014,34(1):1-6.ZHANG Q,ZENG L H,CAI Y L,et al.Genetic analyses on growth and form traits of open-pollinated families of Cinnamomum camphora[J].J Cent South Univ For Technol,2014,34(1):1-6.DOI: 10.14067/j.cnki.1673-923x.2014.01.008.
[24]梁德洋,金允哲,赵光浩,等.50个红松无性系生长与木材性状变异研究[J].北京林业大学学报,2016,38(6):51-59.LIANG D Y,JIN Y Z,ZHAO G H,et al.Variance analyses of growth and wood characteristics of 50 Pinus koraiensis clones[J].J Beijing For Univ,2016,38(6):51-59.DOI: 10.13332/j.1000-1522.20150465.
[25]DIAO S,HOU Y M,XIE Y H,et al.Age trends of genetic parameters,early selection and family by site interactions for growth traits in Larix kaempferi open-pollinated families[J].BMC Genet,2016,17(1):104.DOI: 10.1186/s12863-016-0400-7.
[26]李安琪,严春晓,连勇机,等.黑木相思半同胞子代家系苗期性状的遗传分析[J].西南林业大学学报(自然科学),2023,43(2):9-16.LI A Q,YAN C X,LIAN Y J,et al.Genetic analysis for seedling traits of half-sib progeny in Acacia melanoxylon[J].J Southwest For Univ (Nat Sci),2023,43(2):9-16.
[27]唐良民,周衍斌.马尾松主要生长性状遗传参数和选择效果分析[J].南方林业科学,2020,48(6):29-34.TANG L M,ZHOU Y B.Analysis on genetic parameters and selection effects of main growth traits in Pinus massoniana[J].South China For Sci,2020,48(6):29-34.DOI: 10.16259/j.cnki.36-1342/s.2020.06.007.
[28]刘宇,徐焕文,李志新,等.白桦杂交子代家系生长变异及稳定性分析[J].植物研究,2015,35(6):937-944.LIU Y,XU H W,LI Z X,et al.Growth variation and stability analysis of birch crossbreed families[J].Bull Bot Res,2015,35(6):937-944.DOI: 10.7525/j.issn.1673-5102.2015.06.023.
[29]张磊.杂种落叶松家系稳定性及优良家系初步选择研究[D].哈尔滨:东北林业大学,2011.ZHANG L.Study on stability of families and selection of superior families of hybrid larch[D].Harbin:Northeast Forestry University,2011.
[30]李火根,黄敏仁,潘惠新,等.美洲黑杨新无性系生长遗传稳定性分析[J].东北林业大学学报,1997,25(6):1-5.LI H G,HUANG M R,PAN H X,et al.The genetic stability analysis of growth for new cottonwood clones[J].J Northeast For Univer,1997,25(6):1-5.
[31]孙英豪.5年生长白落叶松高生长遗传变异与多点稳定性分析[D].哈尔滨:东北林业大学,2018.SUN Y H.Genetic variation and multipoint stability analysis of high growth of Larix olgensis henry grown in 5 years[D].Harbin:Northeast Forestry University,2018.
[32]刘宇,徐焕文,尚福强,等.3个地点白桦种源试验生长稳定性分析[J].北京林业大学学报,2016,38(5):50-57.LIU Y,XU H W,SHANG F Q,et al.Growth stability of Betula platyphylla provenances from three sites[J].J Beijing For Univ,2016,38(5):50-57.DOI: 10.13332/j.1000-1522.20150463.
[33]李红盛,汪阳东,徐刚标,等.山苍子家系幼林生长性状遗传变异及稳定性分析[J].林业科学研究,2018,31(5):168-175.LI H S,WANG Y D,XU G B,et al.Analysis of the genetic variation and stability of growth traits of Litsea cubeba young plantation[J].For Res,2018,31(5):168-175.DOI: 10.13275/j.cnki.lykxyj.2018.05.23.
[34]YUAN C Z,ZHANG Z,JIN G Q,et al.Genetic parameters and genotype by environment interactions influencing growth and productivity in Masson pine in east and central China[J].For Ecol Manag,2021,487:118991.DOI: 10.1016/j.foreco.2021.118991.
(责任编辑 郑琰燚)
基金项目:国家重点研发计划(2022YFD220030202,2022YFD2201002)。
第一作者:王佳兴(wjx0707@yeah.net),博士生。*通信作者:张含国(hanguozhang1@sina.com),教授。
引文格式:王佳兴,闫平玉,孙佰飞,等.长白落叶松自由授粉家系生长变异及优良家系早期选择[J]. 南京林业大学学报(自然科学版),2024,48(5):81-89.
WANG J X,YAN P Y,SUN B F,et al.Growth variation and superior families early selection of Larix olgensis free-pollinated families[J]. Journal of Nanjing Forestry University (Natural Sciences Edition),2024,48(5):81-89.