多倍体水稻DHF-CBS基因的抗旱性鉴定
2024-10-09葛杰何玉池展帆李之龙




摘要:为探究干旱胁迫下DHF-CBS基因调控多倍体水稻(Oryza sativa L.)的抗旱机制,选取2种水稻品种,分别为多倍体植株中花-11(DHF-CBS基因)、突变体植株UC-32(不表达DHF-CBS基因),对2种水稻的GUS化学染色结果、组织含水量、脱落酸(ABA)含量、硫代巴比妥酸活性物质(TBARS)含量、谷胱甘肽(GSH)含量等进行分析。结果表明,干旱胁迫第7天,中花-11仍有部分叶片保持绿色且坚挺,干旱胁迫第11天时整株叶片变黄,甚至有的叶片变枯,经过干旱胁迫后的表型观察发现,中花-11比UC-32的抗旱能力强;对水稻的含水量、ABA含量、TBARS含量、GSH含量进行分析,表明中花-11的抗旱性优于UC-32,说明DHF-CBS基因增强了水稻的抗旱能力。
关键词:多倍体;水稻(Oryza sativa L.);DHF-CBS基因;抗旱性
中图分类号:S511 文献标识码:A
文章编号:0439-8114(2024)09-0216-05
DOI:10.14088/j.cnki.issn0439-8114.2024.09.036 开放科学(资源服务)标识码(OSID):
Identification of drought resistance of polyploid Oryza sativa L. DHF-CBS gene
GE Jie1, HE Yu-chi2, ZHAN Fan1, LI Zhi-long1
(1. College of Life Sciences/Key Laboratory of Agricultural Biotechnology, Xinjiang Agricultural University, Urumqi 830052, China;
2. School of Life Sciences, Hubei University, Wuhan 430062, China)
Abstract: In order to explore the drought resistance mechanism of DHF-CBS gene regulation in polyploid Oryza sativa L. under drought stress, two varieties of Oryza sativa L. were selected, namely polyploid plant Zhonghua-11 (DHF-CBS gene) and mutant plant UC-32 (non expressing DHF-CBS gene),the GUS chemical staining results, tissue moisture content, abscisic acid (ABA) content, thiobarbituric acid reactive substance (TBARS) content, glutathione (GSH) content, etc. of two types of Oryza sativa L. were analyzed. The results showed that on the 7th day of drought stress, some leaves of Zhonghua-11 remained green and firm. On the 11th day of drought stress, the entire plant’s leaves turned yellow, and some leaves even withered,after observing the phenotype under drought stress, it was found that Zhonghua-11 had stronger drought resistance than UC-32. Analysis of the water content, ABA content, TBARS content, and GSH content of Oryza sativa L. showed that the drought resistance of Zhonghua-11 was better than that of UC-32, indicating that the DHF-CBS gene enhanced the drought resistance of Oryza sativa L..
Key words: polyploid; Oryza sativa L.; DHF-CBS gene; drought resistance
DHF-CBS基因(Drought high fertility CBS)是CBS家族的重要成员,含有2个CBS结构域及特有的PBI结构,CBS结构域常作为参与细胞能量代谢的传感器发挥作用,能够对细胞内的氧化还原平衡起到维持和调节作用,也可能在植物抗逆性和植物生殖发育过程中发挥重要作用。但是关于干旱胁迫下DHF-CBS基因调控多倍体水稻(Oryza sativa L.)生殖期抗旱的机制鲜有报道。DHF-CBS基因与非生物胁迫、花粉和胚特异表达相关,可能参与开花期抗旱性调控[1]。前期比较蛋白质组学和基因芯片数据分析均表明,CBS家族成员DHF-CBS基因在花药中显著高表达,GUS染色和定量PCR均证实其在花药中的优势表达。干旱胁迫下DHF-CBS基因花粉育性和结实率均显著下降,基因功能补偿后恢复正常,表明该基因可能参与生殖阶段抗旱性调控[2]。因此,开展DHF-CBS基因对多倍体水稻花期抗旱性的调控研究具有一定的理论和实践意义。
干旱胁迫严重影响作物的生长、细胞代谢和作物产量。粮食作物受到干旱的影响导致大幅减产,对全世界的粮食安全造成威胁。植物对干旱的适应性包含诱发防御基因表达、抑制发育相关基因、促进气孔关闭、抑制细胞分裂和分化等[3]。多倍体水稻相对二倍体水稻基因组倍性增加,不仅使抗性基因的数量增加、遗传变异范围变广,而且提高了优良基因组合的概率,增强水稻对各种极端环境的适应能力,是抗旱研究的良好材料。水稻在不同发育阶段对干旱胁迫的应激响应方式和程度有很大不同。水稻生殖期是对干旱最敏感的时期,受干旱影响最大的是水稻花粉发育,干旱导致大量花粉发育不正常,最终导致不育,这种不可逆变化严重影响水稻产量。产量是评价水稻抗旱性的关键指标,开花阶段的抗旱性和产量形成直接相关。多倍体化是推动植物进化和新品种形成的重要因素,多倍体植物较二倍体植物表现出更高的环境适应能力。多倍体植物气孔变大、密度变小、表皮加厚,这些特征减少了水分流失,提高了抗旱性。
1 材料与方法
1.1 材料
水稻品种为多倍体植株中花-11(DHF-CBS基因)、突变体植株UC-32(不表达DHF-CBS基因),均由湖北大学提供,盆栽种植于新疆农业大学。
1.2 方法
1.2.1 水稻生殖期干旱胁迫 挑选健康、子粒饱满的水稻种子,催芽48 h,种子露白后,选取长势相近且发芽生长良好的种子,放于人工气候箱中,光照时间∶黑暗时间=16 h∶8 h。种子成苗后,移栽到温室土壤中继续培养。 待其生长至生殖期,对其进行干旱胁迫,安排3个重复。
1.2.2 GUS染色检测基因DHF-CBS启动子的活性 为确保其准确性,对中花-11和UC-32进行GUS染色,检测DHF-CBS基因的表达情况,基因的启动子位于5′端,处于基因转录起始位点的上游,参与基因的一系列表达和调控,可以被RNA聚合酶识别和结合,是重要的顺式元件。
1.2.3 植物相对含水量 植物组织含水量反映植物组织的水分生理状况[4],测定水稻根、茎、叶组织的相对含水量可以明确不同品种间水稻的抗旱差异。水在作物生长发育中发挥着重要作用,水参与各种代谢过程[5],其含量直接影响作物的新陈代谢速度。植物组织中的水分以2种状态存在:一种是与原生质胶体紧密结合的结合水,另一种是可以自由移动的游离水。游离水和结合水的含量与植物的生长和抗旱性密切相关。当游离水与结合水的含量较高时,植物组织或器官的代谢活性通常更旺盛,生长也更快;当游离水与结合水的含量较低时,植物生长速度较慢 [6]。因此,游离水和结合水的含量可以作为植物组织代谢活性和抗逆性的重要生理指标。一般情况下,作物生长旺盛的部位(如根尖、绿叶、幼苗等)的含水量较高,为60%~90%;在休眠期作物的含水量相对较少,约为40%,干燥种子中含水量最少,约为10%[7]。作物直接从土壤中吸收水分,土壤含水量是直接影响作物根系吸水的重要因素。
1.2.4 脱落酸含量 脱落酸(ABA)是植物体内产生的一种抑制植物生长的应激激素,它能够促使植物气孔关闭,降低蒸腾作用,促进植物组织叶片脱落[8]。在缺水条件下,植物叶片中ABA含量增加,导致气孔关闭。这是因为ABA促进钾离子、氯离子和苹果酸离子的流出,从而促进气孔关闭。在植物叶片上喷洒ABA水溶液可以关闭气孔,降低蒸腾速率。因此,ABA可以作为一种抗蒸腾剂。一般来说,干旱、寒冷、高温、盐度和内涝等不利条件可以迅速使植物体内的ABA含量增加,增强其抗逆性。ABA能显著减少高温对叶绿体超微结构的损伤,提高叶绿体的热稳定性;ABA可以诱导某些酶的生物合成,提高植物的抗寒性、耐涝性和耐盐性。ABA也可以作为植物抵御盐、热和冷损伤的物质,这可能与其促进植物产生新的应激蛋白有关。
1.2.5 硫代巴比妥酸活性物质 脂质过氧化是植物和动物细胞损伤的一种公认机制,被用作细胞和组织氧化应激的指标。活性氧与生物膜的磷脂、酶和膜受体相关的多不饱和脂肪酸侧链及核酸等大分子物质反应形成脂质过氧化产物,如丙二醛(MDA)[9]。MDA可通过与硫代巴比妥酸的受控反应进行定量,生成硫代巴比妥酸活性物质(TBARS)[10]。
1.2.6 谷胱甘肽含量 DHF-CBS基因是调控谷胱甘肽(GSH)酶的重要影响因子,DHF-CBS是编码同型半胱氨酸转化为胱硫醚的关键基因。谷胱甘肽酶功能缺失会导致同型半胱氨酸含量升高,造成不可逆的DNA损伤,同时阻碍下游谷胱甘肽的合成,抗氧化能力下降[11]。胱硫醚β-合成酶是同型半胱氨酸与丝氨酸缩合形成胱硫醚的关键酶,胱硫醚β-合成酶在裂解酶的作用下形成半胱氨酸和α-酮丁酸,最后半胱氨酸经过后续转化形成抗氧化的小分子谷胱甘肽[12]。胱硫醚β-合成酶的功能缺失导致同型半胱氨酸代谢受阻,一方面高同型半胱氨酸易使DNA产生不易修复的损伤,另一方面,下游抗氧化物谷胱甘肽的合成受阻,造成过氧化物及自由基的过量产生[13]。谷胱甘肽是一种抗氧化剂,具有增强生物抗氧化功能、促进生长等功能。作为一种抗氧化剂,它可以帮助生物体消除自由基和过氧化物,从而保持生物体的相对稳定。
谷胱甘肽具有抗氧化、抗应激及促进碳水化合物、脂肪和蛋白质代谢的作用。谷胱甘肽调节植物中各种酶的活性,增强作物对寒冷、干旱、干热和其他胁迫的抵抗力[14,15]。氧化型谷胱甘肽也可以在植物中转化为还原型谷胱甘肽,具有提高作物抗逆性的作用。谷胱甘肽具有促进生长的作用,可以增强生物体对不良环境的抵抗力。植物在胁迫条件下可能会增加体内活性氧的含量,从而发生植物的氧化应激反应。为了避免对自身组织和细胞的氧化损伤,植物会在活性氧含量超过一定阈值后激活其抗氧化系统,从而清除体内大部分多余的活性氧,并确保其在氧化和还原过程中的稳态[16]。谷胱甘肽帮助植物保持正常的免疫系统功能,具有抗氧化和整合解毒功能。谷胱甘肽能参与到生物体的转化作用中,把有害物质转化为无害物质,故测定其生理指标对DHF-CBS功能鉴定有重要意义。
2 结果与分析
2.1 DHF-CBS基因影响水稻抗旱特性
由图1可知,在干旱胁迫第3天时UC-32叶片出现卷曲、叶片末端发黄,干旱胁迫第7天,整株叶片变黄,甚至有的叶片变枯。
由图2可知,干旱胁迫第7天,中花-11仍有部分叶片保持绿色且坚挺,干旱胁迫第11天时整株叶片变黄,甚至有的叶片变枯。经过干旱胁迫后的表型观察发现,中花-11比UC-32的抗旱能力强,说明DHF-CBS基因增强了水稻的抗旱能力。
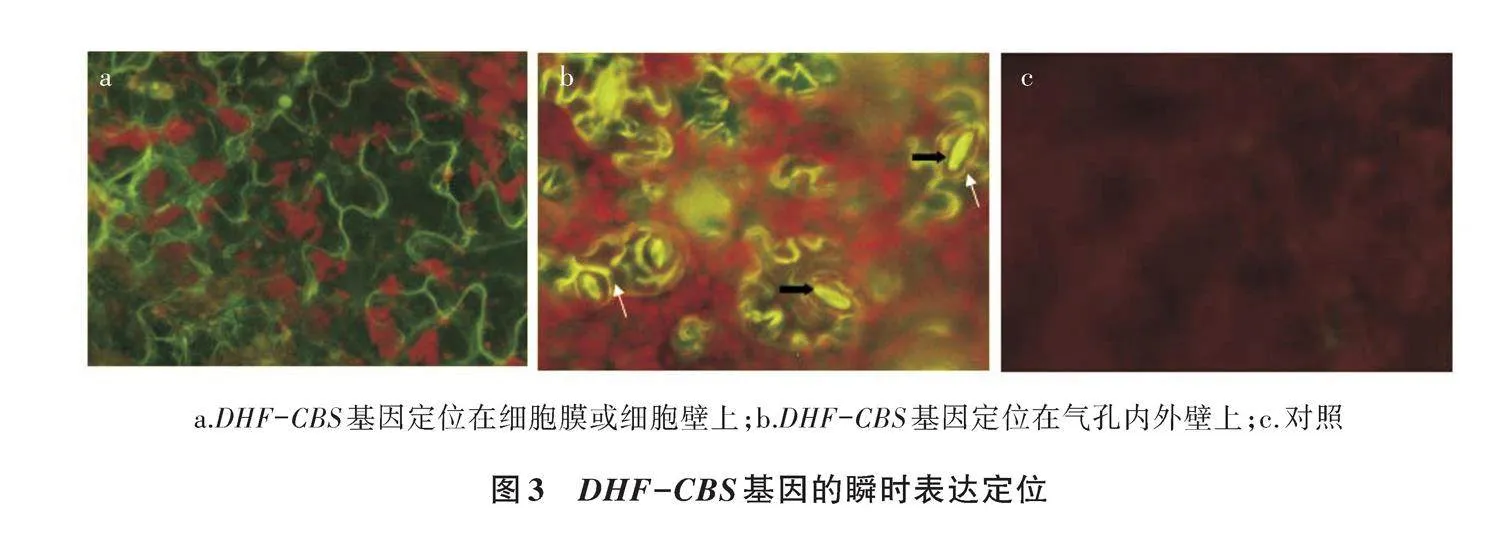
2.2 DHF-CBS基因的瞬时定位表达
研究DHF-CBS基因的亚细胞定位,构建瞬时表达载体。质粒转化农杆菌,暗培养48 h,在Olympus BX51荧光显微镜下观察。结果(图3)表明,约80%的烟草叶片发出GFP的绿色荧光,植株细胞质膜和叶脉都发出绿色荧光,烟草气孔内壁发出较强的绿色荧光,表明DHF-CBS蛋白是参与组成细胞膜或细胞壁的蛋白。DHF-CBS基因在气孔内壁有较强表达,与水分调控关系比较密切。
2.3 GUS染色结果分析
由于绝大多数植物细胞不具有内源β-葡萄糖苷酶(GUS)活性,因此GUS基因被广泛用作转基因植物的报告基因。GUS可以水解X-Gluc形成蓝色物质。初始产物是无色吲哚衍生物,没有颜色,通过氧化二聚化形成5,5’-二溴-4,4’-二氯靛蓝染料。对试验材料的整花、种子及叶片前、中、末段进行GUS染色,染色结果(图4)可知,UC-32各部位均未显蓝色。而中花-11各部位都有蓝色显现,在花萼、柱头、花丝、花药中蓝色明显,表达量较高;在种子和叶片上也观察到明显的蓝色物质;在叶片前端幼嫩叶片部位有蓝色点状分布,表明GUS基因表达量较高,叶片中部GUS基因表达量较少。DHF-CBS基因在水稻中花-11中的不同组织部位存在特异性表达,水稻花中表达量较高。
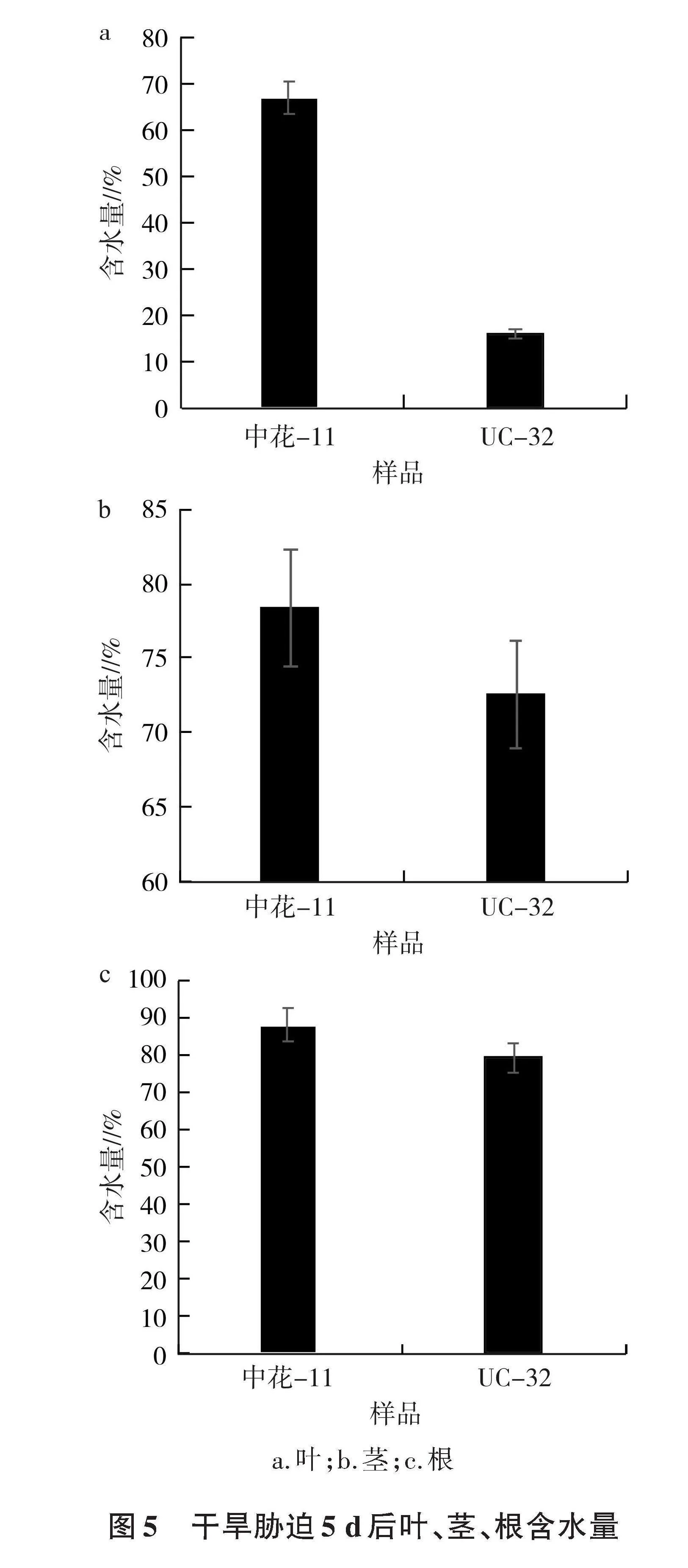
2.4 组织含水量分析
对中花-11和UC-32进行干旱胁迫处理,5 d后分别测定水稻叶、茎、根组织的含水量,重复5次并取平均值。由图5可知,中花-11叶、茎、根组织的含水量均高于UC-32,表明中花-11的抗旱性优于UC-32。
2.5 ABA含量分析
分别在1、3、6、24 h时测定受到干旱胁迫和没有受到干旱胁迫的水稻ABA含量,由图6可知,24 h时,没有受到干旱胁迫的水稻,ABA含量均明显低于干旱胁迫。UC-32的ABA含量上升幅度明显大于中花-11,可能是一种抗逆保护机制,在缺水条件下,植物叶片中ABA含量增加,导致气孔关闭。这是因为ABA促进钾离子、氯离子和苹果酸离子的流出,从而促进气孔关闭,降低蒸腾速率。水稻通过增加ABA含量抑制叶片气孔的开放,促进外部营养器官的脱落,减少水稻在干旱胁迫下对营养分配的压力,把大部分的营养物质和促生长因子集中分配给根和茎。
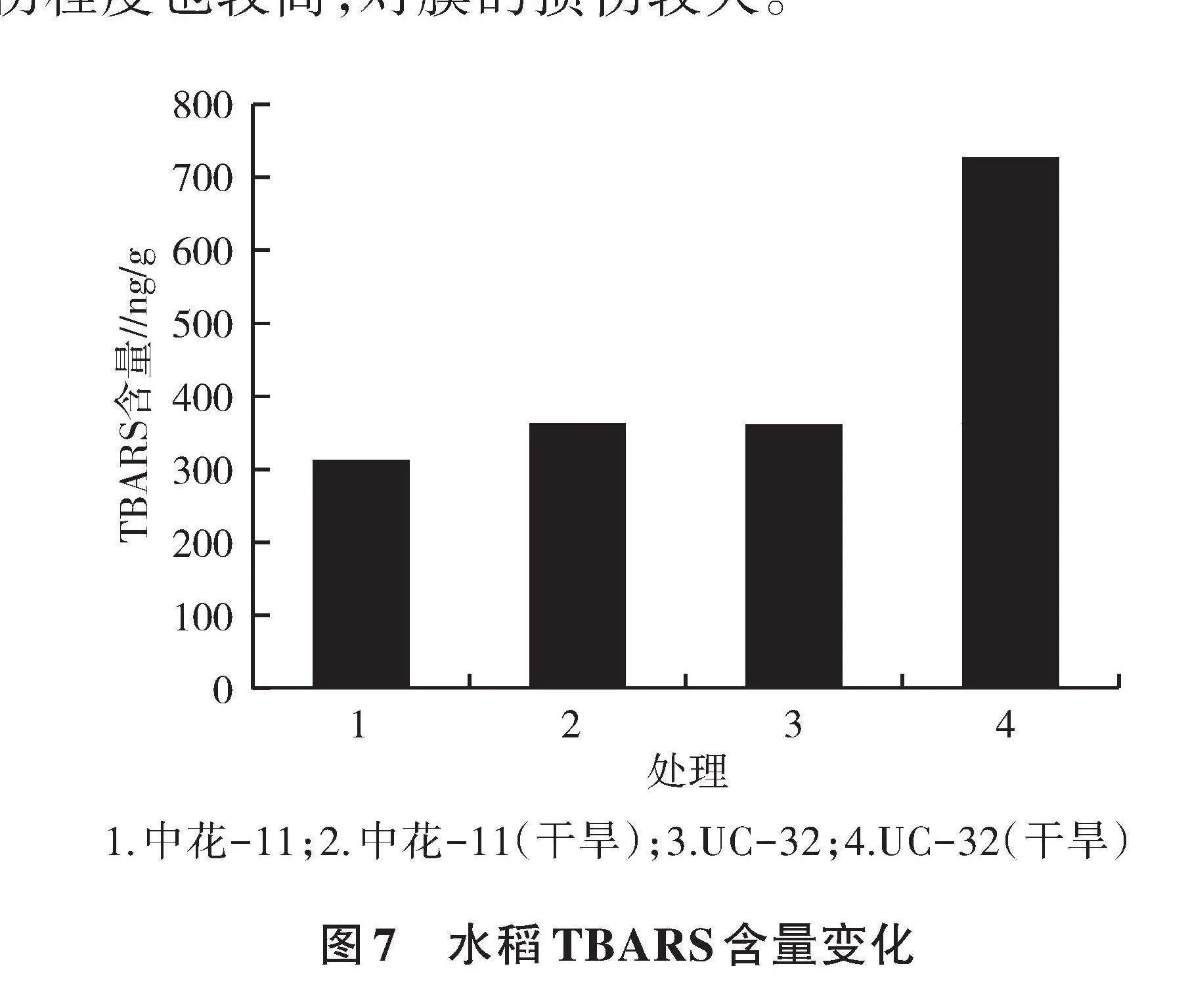
2.6 TBARS含量分析
由图7可知,在没有干旱胁迫下,UC-32的TBARS含量略高于中花-11。在干旱胁迫5 d后,中花-11和UC-32的TBARS含量均升高,其中,UC-32的增加幅度明显高于中花-11,说明在干旱胁迫下,UC-32的脂质过氧化程度高于中花-11,其氧化损伤程度也较高,对膜的损伤较大。
1.中花-11;2.中花-11(干旱);3.UC-32;4.UC-32(干旱)
2.7 GSH含量分析
由图8可知,在没有干旱胁迫下,中花-11的GSH含量明显高于UC-32。在干旱胁迫5 d后,中花-11和UC-32的GSH含量均升高,其中,UC-32升高的幅度明显高于中花-11,结果表明,中花-11对GSH的需求量更小,也从侧面反映出UC-32的抗旱能力低于中花-11。
3 小结
抗旱是一个综合的生理过程,与品种抗旱强度有关的因素很多,本研究只探讨了DHF-CBS基因与水稻抗旱性的关系,UC-32的GSH含量整体低于中花-11的原因需要进一步研究。干旱胁迫第7天,中花-11仍有部分叶片保持绿色且坚挺,干旱胁迫第11天时整株叶片变黄,甚至有的叶片变枯,经过干旱胁迫后的表型观察发现,中花-11比UC-32的抗旱能力强;对水稻的含水量、ABA含量、TBARS含量、GSH含量进行分析,表明中花-11的抗旱性优于UC-32,说明DHF-CBS基因增强了水稻的抗旱能力,本研究为进一步解析DHF-CBS基因在水稻抗旱中的作用机制提供参考。
参考文献:
[1] 余 欣, 童 飞, 詹 妮,等. 干旱-高温交叉胁迫对水稻幼苗光合特性的影响[J].干旱地区农业研究,2022,40(3):72-78.
[2] 段 梅.水稻OsI2基因的功能鉴定[D].武汉:华中农业大学,2009.
[3] 栗雨勤,张文英,王有增,等.作物抗旱性鉴定指标研究及进展[J].河北农业科学,2004,8 (1):58-61.
[4] 孙彩霞,沈秀瑛,刘志刚.作物抗旱性生理生化机制的研究现状和进展[J].杂粮作物,2002, 22(5):285-288.
[5] 穆 平,李自超,春 平,等.水、旱稻根系性状与抗旱性相关分析及其QTL定位[J].科学通报, 2003,48(20):2162-2169.
[6] BOXALL S F, KADU N, DEVER L V, et al.Kalancho? PPC1 is essential for crassulacean acid metabolism and the regulation of core circadian clock and guard cell signaling genes[J]. Plant cell, 2020,32:1136-1160.
[7] 高越峰,荆玉样,沈世华,等.高赖氨酸蛋白基因导入水稻及可育转基因植株的获得[J].植物学报,2001,43(5):506-511.
[8] 刘 琴,孙 辉,何道文.干旱和高温对植物胁迫效应的研究进展[J].西华师范大学学报(自然科学版),2005(4):364-368.
[9] 孟宪梅,黄义德,李奕松,等.水稻若干生理指标与品种抗旱性关系的研究[J].安徽农业大学学报,2003(1):15-22.
[10] 王雪珂. 水稻抗旱性鉴定墒情智能控制模型的研究与应用[D]. 合肥: 安徽农业大学, 2023.
[11] 王贺正,马 均,刘慧远.水稻抗旱性研究现状与展望[J].中国农学通报,2005,21(1):111-113.
[12] 赵凤云,张 慧.耐非生物胁迫转基因水稻的培育——现在和未来[J].生物工程学报,2007,23(1)1-6.
[13] 罗利军,张启发.栽培稻抗旱性研究的现状与策略[J].中国水稻科学,2001,15(3):209-214.
[14] PELLINEN R, PALVA T, KANGASJRVI J. Subcellular localization of ozone-induced hydrogen peroxide production in birch (Betula pendula) leaf cells[J]. Plant journal, 2010, 20(3):82-87.
[15] 刘菊红.水稻CONSTANS-like基因Ghd2调控抗旱性和衰老的功能研究[D].武汉:华中农业大学,2016.
[16] ALBACETE A A,MARTINEZ-ANDUJAR C,PEREZ A F.Hormonal and metabolic regulation of source-sink relations under salinity and dr ought: From plant survival to crop yield stability[J].Biotechnology advances,2014,32:12-30.
收稿日期:2023-04-17
基金项目:国家自然科学基金项目(32160494);新疆农业大学大学生创新创业训练计划项目(dxscx2022068)
作者简介:葛 杰(1978-),女,新疆乌鲁木齐人,副教授,博士,主要从事植物抗逆分子生物学研究,(电话)17799659806(电子信箱)29257741@qq.com。
葛 杰,何玉池,展 帆,等. 多倍体水稻DHF-CBS基因的抗旱性鉴定[J]. 湖北农业科学,2024,63(9):216-220.
