哀牢山植物群落数量分类与排序
2024-10-08徐乔芬欧光龙付磊
摘要为了明确影响哀牢山植物群落分布的主要环境因子,本研究采用双向指示种分析(TWINSPAN)和典范对应分析(CCA),对研究区45个样方分布与环境因子之间的生态关系进行数量分类与排序。结果表明,TWINSPAN分类将45个样地点中170个物种划分为7个群丛,其中群丛Ⅲ的物种多样性最丰富,群丛Ⅴ物种多样性最单一,仅包括11和20号样地中的18个物种;CCA排序结果表明,海拔是哀牢山植物群落分布的主要影响因素,其次是孔隙度和温度,土壤有机质和土壤全磷对植物群落分布影响不明显。哀牢山植物群落数量分类变化的分析对揭示植物群落空间分布特征和植物种质资源调查及其保护具有重要的意义。
关键词植物群落;分类;排序;双向指示种分析;典范对应分析
中图分类号S718.54文献标识码A文章编号1007-7731(2024)18-0049-05
DOI号10.16377/j.cnki.issn.1007-7731.2024.18.011
Quantitative classification and ranking of plant communities in Ailao Mountain
XU Qiaofen1OU Guanglong1FU Lei2
(1Southwest Forestry University, Kunming 650224, China;
2Kunming Forest Resources Management Station, Kunming 650221, China)
Abstract In order to determine the major environmental factors affecting plant community distribution in Ailao Mountain, two-way indicator species analysis (TWINSPAN) and canonical correspondence analysis (CCA) were used to quantitatively classify and sequence the ecological relationships between 45 communities and environmental factors in study area. The results showed that, 170 species from 45 quadrats were divided into 7 clusters by TWINSPAN classification. Cluster Ⅲ had the most abundant species diversity, while cluster Ⅴ had the most unitary species diversity, including only 18 species from plots 11 and 20. The results of CCA sequencing showed that, elevation was the main factor affecting the distribution of plant community in Ailao Mountain, followed by porosity and temperature. Soil or2vR32N24wCaM1z7wXlkGyg==ganic matter and soil total phosphorus had no significant effect on the distribution of plant community. The analysis of the change of plant community quantity classification in Ailao Mountain is of great significance for revealing the spatial distribution characteristics of plant community and the investigation and protection of plant germplasm resources.
Keywords plant communities; classification; ranking; two-way indicator species analysis; canonical correspondence analysis
云南哀牢山属横断山脉南段中山峡谷亚区,地处横断山系和云南高原两大自然地理区域相结合的部位,是云南省的主要生态屏障之一。哀牢山作为亚热带南、北部的过渡地带是地理分布东西交汇、南北过渡之地。哀牢山分布着以云南特有植物为优势物种的亚热带常绿阔叶林,保持了生态系统的原真性和完整性,为该地区提供了安全生态屏障,以及为公众提供优质的生态服务。
植物群落数量分类与排序方法可以整体研究植物群落分类及其内部特征,是认识某一区域内植被特征的重要方法之一,是进行植物群落多样性研究的基础[1-3]。该方法可以较为准确、客观地揭示植物群落特征与生态环境之间的关系,是研究植物生态学的重要途径之一[4]。双向指示种分析(Two-way indicator species analysis,TWINSPAN)是用数学方法来完成分类过程,以揭示生态关系,其分类的结果能反映一定的生态规律;除趋势对应分析(Detrended correspondence analysis,DCA)和典范对应分析(Canonical correspondence analysis,CCA)是研究植被连续变化的方法,能够解释植被或植物种的分布与环境之间的关系[5-6]。在植被数量研究中,TWINSPAN分类与DCA、CCA排序的结合分析能够更好地分析物种分布格局与环境之间的关系[7]。本研究采用TWINSPAN和CCA对哀牢山自然保护区的植物群落进行分类排序研究,较为系统地分析环境因子对植物群落组成和分布规律的影响,为今后哀牢山植物群落多样性的研究和植物群落演替的预测提供参考。
1 材料与方法
1.1 研究方法
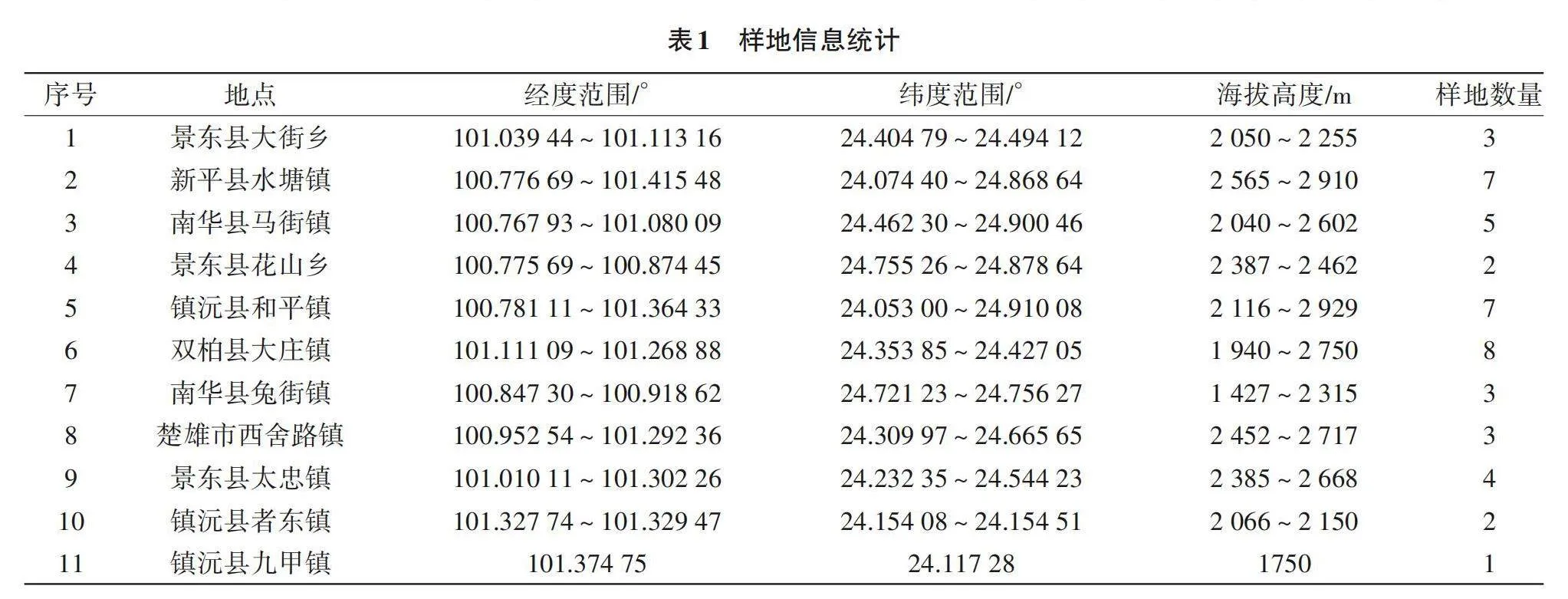
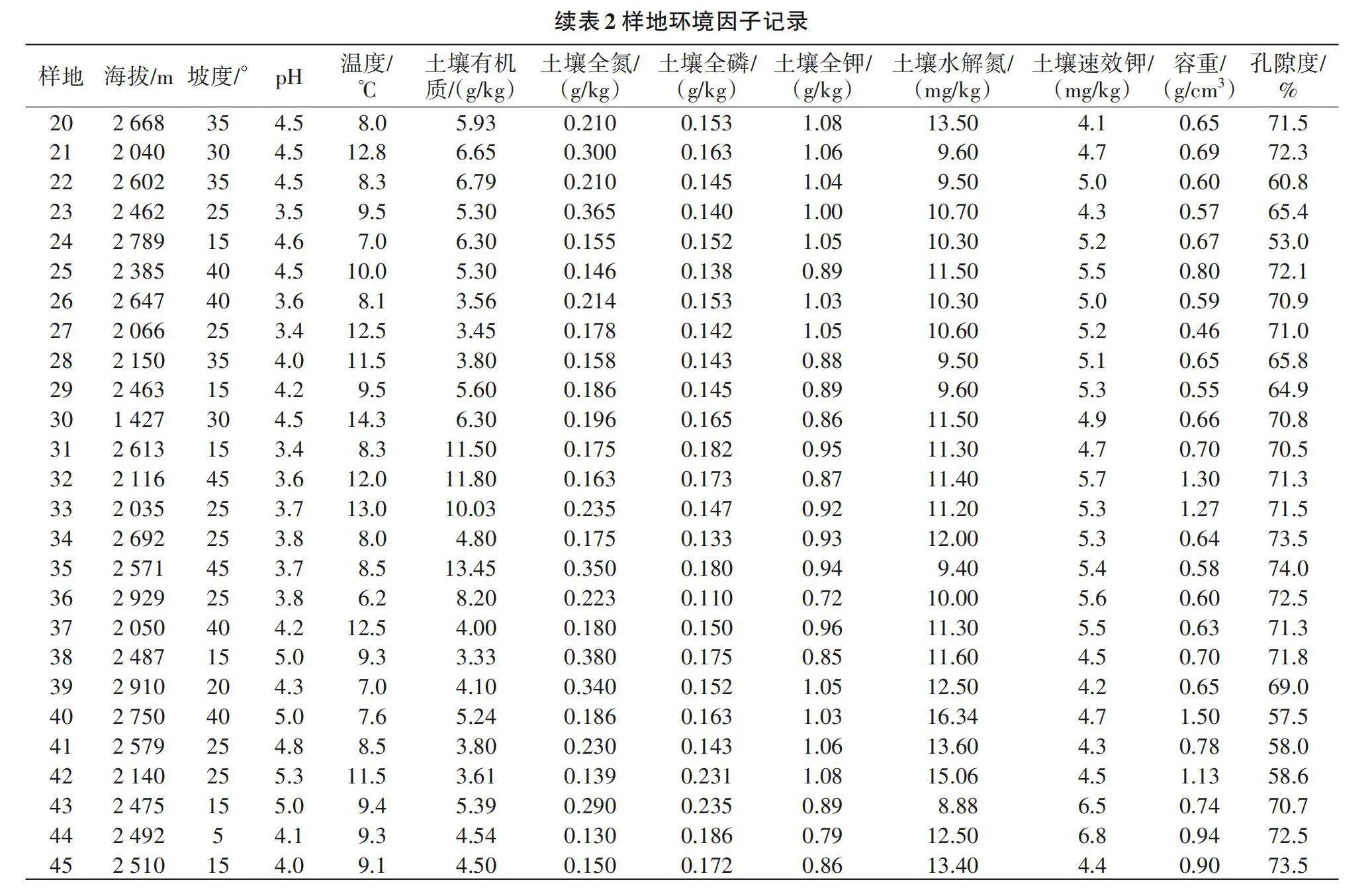
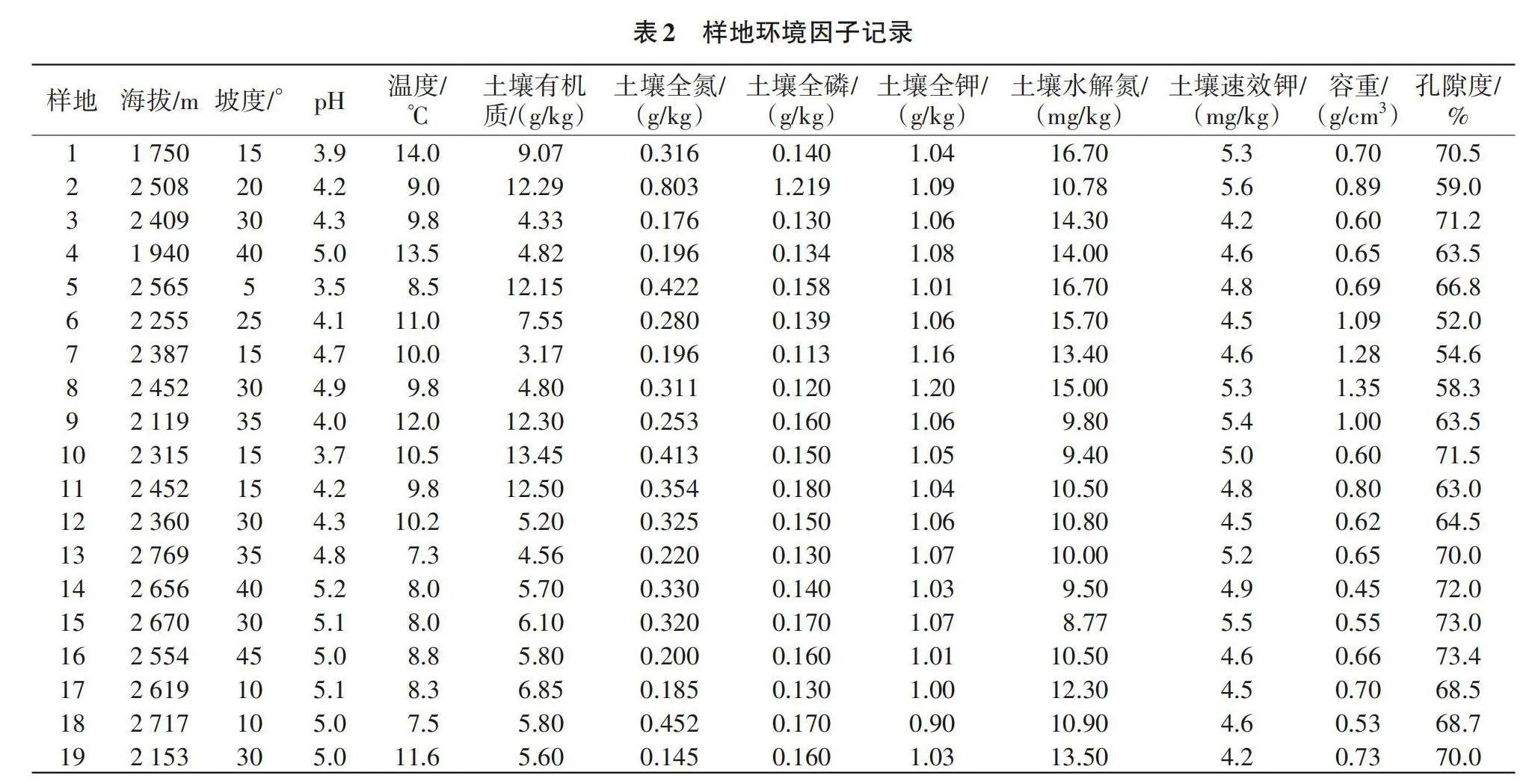
采用样线法和典型样地法[8]对研究区内的植物物种和植被进行全面调查,选择样地时尽量避开人为干扰的地段,以海拔为梯度,选取研究区垂直地带不同植被类型和植物群落。由表1可知,试验共设置45个样方,每隔200 m海拔设置一块样地,乔木层选择大小20 m×20 m的样方,记录乔木层树种种类、株数、树高、胸径和郁闭度等(高度>5 m起测);在灌木层和草本层分别设置4块5 m×5 m小样方,记录种类、株(丛)数和树高等。记录样地的基本情况,如小地名、群落名称、平均高度、郁闭度、干扰情况、地形(海拔、坡向和坡度等)、土壤(养分、pH和物理性质等)和气候(温度、湿度等),如表2所示。
1.2 记录指标和方法
1.2.1 重要值计算 重要值是表示物种在群落中优势度的一个指标,可以表示不同物种在群落中的功能地位[9]。其作为TWINSPAN植物群落分类的基础数据,被广泛用于植被分类研究[10-12]。其作为综合指标反映物种的基本特征[13],计算公式如式(1)~(2)。
乔木重要值=(相对多度+相对优势度+相对高度)/3 (1)
灌木、草本重要值=(相对盖度+相对高度)/2 (2)
1.2.2 数量分类 为了消除稀有种对分类结果的影响,在对哀牢山植物群落进行TWINSPAN分类时,将样方内重要值小于0.1且出现1次的植物种类剔除,最后用于TWINSPAN分类的植物种类数为170种,然后将170个植物种的重要值与45个植物群落样方组成的植被矩阵进行分类。
1.2.3 环境因子处理 冗余分析(Redundancy analysis RDA)和CCA是基于对应分析(Correspon-dence analysis, CA)发展而来的一种排序方法,将对应分析与多元回归分析相结合,每一步计算均与环境因子进行回归,环境因子主要参照哀牢山森林生态系统研究中土壤剖面理化性质和土壤养分的含量[14],又称多元直接梯度分析。
1.2.4 CCA排序 选择排序方法前先用丰度矩阵数据做DCA分析,分析结果中排序轴大于4.0,推荐选CCA;排序轴在3.0~4.0,选RDA和CCA均可,小于3.0则推荐使用RDA。经过DCA分析排序轴长度分别为4.07、5.26、5.36和5.63,结果大于4.0,因此选择线性模型中的CCA进行分析。
1.3 数据处理
采用Excel软件分析计算各样地植物群落重要值和丰度;运用PC-ORD 5.0软件进行TWINSPAN植物群落分类;采用CANOCO 5.0软件对样地进行CCA排序分析。
2 结果与分析
2.1 TWINSPAN分类
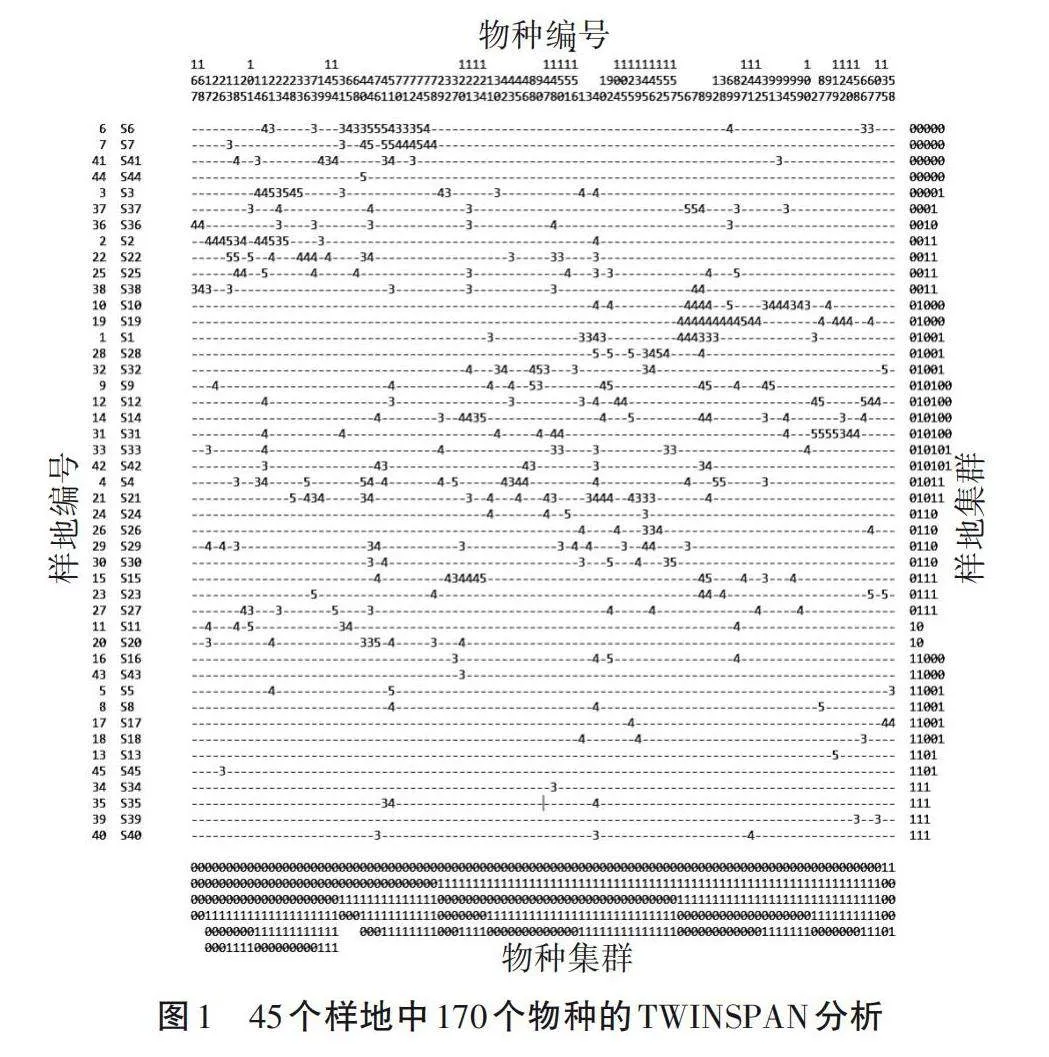
对45个样方中170种植物进行TWINSPAN分析,结果见图1。TWINSPAN双向分类矩阵横排是170个植物种类的编号,竖排是45个样地的编号,以二进制代码表示分类指标的分类水平,样地编号与植物种类交叉处的数量级表示分类指标的相对值。TWINSPAN分析结合群落命名原则与样方实际情况[15],适当进行调整,最终划分了7个群落类型。
旱冬瓜()、珍珠花()和具芒碎米莎草()为群丛Ⅰ,包括3、6、7、37、41和44号样方中的40个物种,旱冬瓜平均高度7.5 m,分布在海拔2 200~2 400 m;灌木层主要是珍珠花,平均高度5 m,伴生种有无柄金丝桃()、云南越橘()等;草本层为具芒碎米莎草,平均高度25 cm,伴生种有野拔子(a)、蓟()、原沼兰()和七小叶崖爬藤()等。
群丛Ⅱ为白穗石栎()、马缨杜鹃()和碎米莎草(),包括2、22、25、36和38号样方中的40个物种,乔木层中白穗石栎平均高度12.3 m,分布在海拔2 550 m左右,华山松()、旱冬瓜也出现在此区域,属于典型的针阔混交林;灌木层主要伴生种有景东十大功劳()、三叶悬钩子()和苍山越橘()等;草本层伴生种有菊三七(Gynura japonica)、宽穗兔儿风()和匍茎沿阶草()等。
石栎()、隐距越橘()和紫茎泽兰()为群丛Ⅲ,包括1、4、9、10、12、14、19、21、28、31、32、33和42号样方中的141个物种,分布于海拔1 750~2 650 m处,乔木优势种石栎平均高度1 m,主要伴生种有高山栲()、瓦山锥()、红花木莲()和旱冬瓜等阔叶树种;林下主要有蒙自连蕊茶()、针齿铁仔()、禾状薹草()和菝葜()等。
中缅木莲()、西南红山茶()、多叶唐松草()和滑竹()为群丛Ⅳ,包括15、23、24、26、27、29和30号样方中的61个物种,分布于海拔1 427~2 670 m,平均高度8 m,此群落植被群落结构较单一,以草本为主,有禾状薹草、菝葜等藤本植物。
元江锥()、露珠杜鹃()和云南越橘为群丛Ⅴ,只有11、20号样方中的18个物种,分布于海拔2 452~2 668 m,此群丛主要为元江栲林、石栎林和露珠杜鹃矮林,平均高度10 m,伴生种有华山松、水红木()、菝葜和光亮玉山竹()等。
疏齿锥()、珍珠花和八月瓜()为群丛Ⅵ,包括5、8、13、16、17、18、43和45号样方中的78个物种,分布于海拔2 510~2 769 m,平均高度11 m,乔木层还有中华木荷()、硬斗石栎();灌木层的伴生种有长尖叶蔷薇()、紫药女贞()、露珠杜鹃和金花小檗()等;草本伴生种有灯心草()、云南莎草()和紫茎泽兰等。
露珠杜鹃和云南兔儿风()为群丛Ⅶ,包括34、35、39和40号样方中的30个物种,此群丛以杜鹃灌丛为主,平均高度3.5 m,散生乔木有云南铁杉()、云南松()、华山松、滇青冈()和杜英();草本伴生种有八月瓜、大八角()、紫茎泽兰和开口箭()等。
2.2 CCA分析
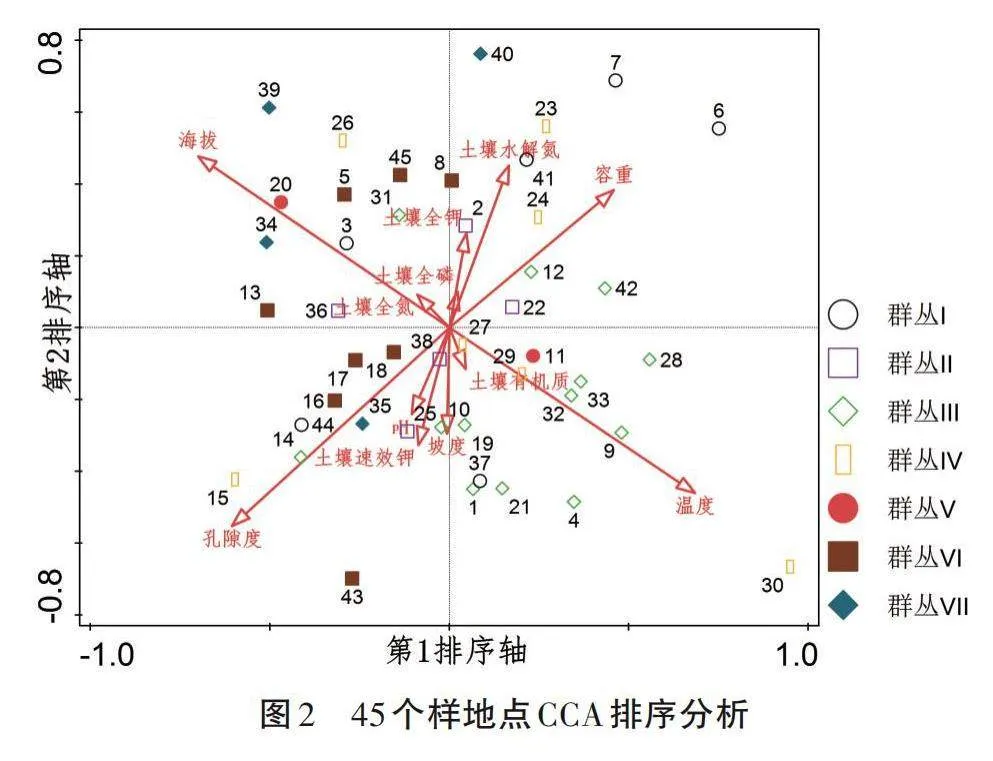
利用环境因子与样地矩阵数据对哀牢山45个样地中的170个植物种类进行CCA排序,分析海拔、坡度、pH、温度和土壤有机质等12个环境因子与7个植被群丛分布的相关关系。CCA排序分析中,箭头所处象限代表环境因子与排序轴间的正负相关性,可根据箭头连线的长度判断某个环境因子与群落分布和种类分布间相关性的强弱,具体表现为距离越近,相关性越大,反之越小。
由图2可知,海拔与群丛Ⅰ中3号样地、群丛Ⅴ中20号样地和群丛Ⅶ中的34号样地相关性较大;孔隙度与群丛Ⅲ中14号样地和群丛Ⅶ中35号样地相关性较大;温度对群丛Ⅲ的影响最大,与群丛Ⅲ大部分样地相关性较大。通过分析可知,海拔、孔隙度、温度、容重和土壤水解氮对哀牢山植物群落的影响较大。
3 结论与讨论
通过TWINSPAN分类和CCA排序对哀牢山植物群落进行数量分类和排序,二者所得结果较为一致,说明这两种方法对哀牢山45个样地点中的170个植物种类在数量分类上的可靠性。根据TWINSPAN分类结果,各个群丛在垂直分布地带上表现为不连续性,有些样地位置相距较远被分成了相同的群丛,群丛分布与研究区域内复杂的地理位置和气候有关。根据CCA分析,影响哀牢山植物群落分布的主要环境因子是海拔,其次是孔隙度和温度,土壤有机质和土壤全磷对植物群落分布影响不大,其原因可能是部分调查样地的植物群落和植被类型较为单一。为了更精准地了解哀牢山的植被分布状况,避免样地布设的主观性,采用多种分析方法探索哀牢山同一区域影响植被分布的主导因子的验证有待进一步开展。
开展哀牢山自然保护区植物群落数量分类与排序研究,进一步掌握植物群落的结构,有助于加强对森林生态系统、生物多样性的保护,提高环境质量,为改善自然保护区周边生态环境,创造开展科研监测工作提供有利条件,为研究区植物群落多样性研究提供参考。
参考文献
[1] 姚帅臣,王景升,丁陆彬,等. 拉萨河谷草地群落的数量分类与排序[J]. 生态学报,2018,38(13):4779-4788.
[2] 陈宝瑞,朱玉霞,张宏斌,等. 呼伦贝尔草甸草原植被的数量分类和排序研究[J]. 武汉植物学研究,2008,26(5):476-481.
[3] 王静娅,王明亮,张凤华. 干旱区典型盐生植物群落下土壤微生物群落特征[J]. 生态学报,2016,36(8):2363-2372.
[4] 苏金娟,刘永萍,刘丽燕,等. 新疆阿勒泰地区典型植物群落数量分类与排序分析[J]. 草业学报,2023,32(9):50-67.
[5] 吴华,张建利,范怡雯,等. 草海流域植物群落结构数量分类与排序[J]. 南京林业大学学报(自然科学版),2013,37(3):47-52.
[6] 苏日古嘎,张金屯,张斌,等. 松山自然保护区森林群落的数量分类和排序[J]. 生态学报,2010,30(10):2621-2629.
[7] 郑东升,刘琪璟. 环境因子对长白山自然保护区森林群落分布的影响[J]. 北京林业大学学报,2023,45(8):57-64.
[8] 方精云,王襄平,沈泽昊,等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性,2009,17(6):533-548.
[9] 欧光龙,彭明春,和兆荣,等. 高黎贡山北段植物群落TWINSPAN数量分类研究[J]. 云南植物研究,2008,30(6):679-687.
[10] 刘秋锋,康慕谊,刘全儒. 中条山混沟地区森林乔木种的数量分类与环境解释[J]. 植物生态学报,2006,30(3):383-391.
[11] 于梦凡. 植物群丛的数量分类方法及对比研究:以辽宁青龙河自然保护区为例[D]. 北京:北京林业大学,2014.
[12] 刘润红,涂洪润,李娇凤,等. 桂林岩溶石山青冈群落数量分类与排序[J]. 生态学报,2019,39(22):8595-8605.
[13] 张钦弟,张金屯,苏日古嘎,等. 庞泉沟自然保护区华北落叶松林的自组织特征映射网络分类与排序[J]. 生态学报,2011,31(11):2990-2998.
[14] 徐跃,张世聪. 哀牢山北段土壤腐殖质组成的垂直分布和腐殖质特性的研究[J]. 生态学杂志,1989,8(2):5-9.
[15] 薛志方,刘彤,王立生,等. 额尔齐斯河流域主要支流平原河谷林群落结构及特征[J]. 植物生态学报,2024,48(3):390-402.
(责任编辑:吴思文)
