普通核桃与深纹核桃中叶绿体基因组密码子使用特征对比
2024-09-24朱艳韩长志













收稿日期:2023-09-06
基金项目:云南省研究生导师团队建设项目(2022100);云南省“兴滇英才支持计划”青年人才专项(YNWR-QNBJ-2020-188)
作者简介:朱 艳(2000-),女,云南昭通人,硕士研究生,主要从事森林保护相关研究。(E-mail)508034093@qq.com
通讯作者:韩长志,(E-mail)swfuhcz@163.com
摘要: 本研究旨在对比分析普通核桃与深纹核桃中叶绿体基因组密码子的使用偏好性,对普通核桃与深纹核桃中各53条蛋白质编码区序列开展预测分析,明确上述2种核桃叶绿体基因组中的最优密码子,为未来开展核桃物种的亲缘关系研究提供理论依据。利用Codon W 1.4.2和在线软件CUSP等分析普通核桃与深纹核桃中的叶绿体基因组密码子,获得相对同义密码子使用度(RSCU)、有效密码子数(ENC)、G+C含量等参数,再进行中性绘图分析、有效密码子数分析及奇偶偏好性分析。普通核桃与深纹核桃中叶绿体基因组的密码子适应指数均为0.167,ENC均在45.00以上,表明密码子偏好性弱。普通核桃与深纹核桃中叶绿体基因组密码子的G+C含量整体表现为:基因中所有密码子第1位碱基G+C的含量(GC1)>基因中所有密码子第2位碱基G+C的含量(GC2)>基因中所有密码子第3位碱基G+C的含量(GC3)。进一步分析发现,第3位碱基的使用频率表现为T大于A,G大于C。从普通核桃中筛选出14个最优密码子,从深纹核桃中筛选出17个最优密码子,其中共有23个最优密码子在第3个位置偏好以A/U结尾。总之,普通核桃和深纹核桃中叶绿体基因组密码子偏好性主要受到自然选择的影响,本研究结果可以为进一步探索核桃抗性遗传基因的改良和表达提供理论基础和数据支撑。
关键词: 普通核桃;深纹核桃;叶绿体;基因组;同义密码子;偏好性
中图分类号: S664.1 文献标识码: A 文章编号: 1000-4440(2024)08-1507-14
Comparison of codon usage characteristics of chloroplast genomes in Juglans regia and Juglans sigillata
ZHU Yan, HAN Changzhi
(College of Forestry, Southwest Forestry University/Key Laboratory of Forest Disaster Warning and Control of Yunnan Province, Kunming 650224, China)
Abstract: The aim of the study was to compare and analyze the usage preference of chloroplast genome codons in Juglans regia and Juglans sigillata, and to predict and analyze 53 protein coding sequences (CDS) in Juglans regia and Juglans sigillata, so as to clarify the optimal codons in the chloroplast genomes of the above two walnuts and provide a theoretical basis for the future research of the genetic relationship of walnut species. Codon W 1.4.2 and online software CUSP were used to analyze the chloroplast genome codons in Juglans regia and Juglans sigillata, and the relative synonymous codon usage (RSCU), effective number of codons (ENC), G+C content and other parameters were obtained. Neutrality plot analysis, effective number of codons analysis (ENC-plot), and parity preference analysis (PR2-plot) were performed. The codon adaptation index of chloroplast genomes in Juglans regia and Juglans sigillata was 0.167, and the ENC was above 45.00, indicating that the codon preference was weak. The G+C content of chloroplast genome codons in Juglans regia and Juglans sigillata was as follows: the content of G+C at the first base of all codons in the gene (GC1)>the content of G+C at the second base of all codons in the gene (GC2)>the content of G+C at the third base of all codons in the gene (GC3). Further analysis showed that the usage frequency of the third base was T>A, G>C. A total of 14 optimal codons were screened in Juglans regia, and 17 optimal codons were screened in Juglans sigillata. Among them, 23 optimal codons preferred the A/U end at the third position. In conclusion, the codon preference of chloroplast genome in Juglans regia and Juglans sigillata is mainly affected by natural selection, which provides theoretical basis and data support for further exploration of the improvement and expression of walnut resistance genes.
Key words: Juglans regia;Juglans sigillata;chloroplast;genome;synonymous codon;bias
生物体内的各种蛋白质由20种氨基酸构成,其中色氨酸、甲硫氨酸只有1个由三核苷酸组成的密码子,其他氨基酸有2~6个由三核苷酸组成的密码子[1]。通过对密码子开展分析能够辅助识别自然界中的遗传信息[2],目前共有64个密码子,18种氨基酸能够利用其简并性实现不同密码子编码同一氨基酸[3]。不同物种、同一物种不同组织或基因中的密码子存在不均衡使用的情况,即同义密码子偏好性(SCUB)[4-6]。最优密码子是在物种高表达基因中频繁出现、使用频率最高的密码子,而部分很少使用甚至不会出现的密码子称为稀有密码子或者非优化密码子[3]。SCUB一般会受到基因突变、自然选择、基因漂移等多种因素影响[7],对其开展研究,有助于了解物种之间的亲缘关系、遗传相似性、基因水平转移、基因分子进化及系统发育等,进而促进外源基因的表达和转基因作物研究的发展[8-10]。叶绿体是绿色植物和藻类能进行光合作用的细胞器[11],具有基因组小、拷贝数多、结构简单、进化保守等诸多优点,因此,叶绿本作为研究对象被广泛应用于DNA条形码分子标记、系统进化以及基因工程等研究领域[8,12]。前人对不同物种叶绿体基因组中SCUB展开了分析工作,如肉苁蓉(Cistanche deserticola Ma)[13]、茶树(Camellia sinensis)[14]、二乔玉兰[Yulania×soulangeana (Soul.-Bod.) D. L. Fu][15]、蒜头果(Malania oleifera Chun & S. K. Lee)[16]、榕树(Ficus microcarpa L. f.)[17]和圆柏(Juniperus chinensis Roxb.)[18]。
作为世界四大干果之一,核桃中含有大量的营养物质[19],自古以来就深受消费者的喜爱[20],同时,作为“木本油料之王”,其在木材建筑、经济效益方面发挥的作用显著[21]。深纹核桃(Juglans sigillata D.)又被称为云南核桃、南方核桃[22],主要分布于中国西南地区和西藏自治区雅鲁藏布江中上游地区,生长于海拔1 300~3 300 m的山坡或山谷林中。普通核桃(Juglans regia L.)是胡桃属植物,主要盛产于中国北方。目前有关上述2种核桃的研究工作主要体现在分子生物学方面,涉及种质资源、表型研究和遗传多样性等[22-24],然而,关于核桃叶绿体DNA的研究较少。本研究拟以深纹核桃、普通核桃中叶绿体基因组的高通量测序结果为基础,通过对比分析其叶绿体基因组蛋白质编码区序列(CDS)的碱基组成,利用中性绘图分析、有效密码子数分析(ENC-plot)及奇偶偏好性分析(PR2-plot)等深入剖析影响密码子偏好性的主要因素,并筛选获得叶绿体基因组的最优密码子,以期为深入开展2种核桃的基因组研究、种质创新研究提供数据支撑。
1 材料与方法
1.1 试验材料
从美国国家生物技术信息中心(NCBI)数据库中获取普通核桃和深纹核桃的叶绿体基因组CDS序列,前者GenBank登录号为KT963008.1,利用EditSeq计算G+C含量为36.23%;后者GenBank登录号为MF167465.1,G+C含量为36.02%。通过在线软件(https://irscope.shinyapps.io/Chloroplot/)绘制普通核桃和深纹核桃的叶绿体基因组图谱。
1.2 试验方法
1.2.1 密码子碱基组成分析 使用Codon W 1.4.2处理数据得到密码子适应指数(CAI)、有效密码子数(ENC)、相对同义密码子使用度(RSCU),通过CUSP分析获得基因中所有NHksyIPAY+9HEPE2JVAsCNJ8waLMkZ+8oAGj/+j4z8Y=密码子第1位碱基、第2位碱基、第3位碱基G+C的含量(GC1、GC2、GC3),最后使用IBM SPSS Statistics、Excel、Origin等分析软件分析并作图[25]。
1.2.2 中性绘图分析 分别以GC12(GC1和GC2的平均值)为纵坐标,GC3为横坐标,绘制散点图,分析密码子使用偏好性的影响因素[26],进而对密码子不同位置的碱基使用性开展相关性分析[27]。
1.2.3 ENC-plot分析 分别以GC3作为X轴变量,ENC作为Y轴变量绘制散点图,通过公式(1)计算出各基因的标准ENC并绘制标准曲线。ENC的理论取值为20~61,其值越接近20表明偏好性越强,反之,其值越接近61表明偏好性越弱,若散点与标准曲线接近,表示密码子使用偏好性的主要影响因素是基因突变,若散点远离标准曲线,说明密码子使用偏好性的主要影响因素是自然选择[28]。为了更好地评估ENC实际观察值与ENC期望值之间的差异,根据公式(2)计算ENC比值并进行分析。
ENC=2+GC3+29/[GC32+(1-GC3)2](1)
ENC比值=(ENCexp-ENCobs)/ENCexp(2)
式中,ENC表示基因中使用有效密码子数;GC3表示基因中所有密码子的第3位碱基G+C的含量;ENCexp表示ENC期望值;ENCobs表示ENC实际观察值。
1.2.4 PR2-plot分析 分别以G3为分子,G3与C3的和为分母,所得比值为X轴变量;以基因中所有密码子第3位碱基A的含量(A3)为分子,A3与基因中所有密码子第3位碱基T的含量(T3)的和为分母,所得比值为Y轴变量,绘制散点图[29]。中心点位置表示A=T、G=C,即密码子不具有偏好性,其他点到中心点的距离表示该基因的偏倚程度。
1.2.5 最优密码子的确定 以密码子的ENC作为分析偏好性的标准,选取升序排列后ENC最低和最高两端各10%的基因,建立基因库。分别计算2组的RSCU值和△RSCU值(高表达基因的RSCU值-低表达基因的RSCU值)[30],确定2种核桃叶绿体基因组的高频密码子(RSCU>1.00)和高表达密码子(△RSCU≥0.08),最优密码子要同时满足RSCU>1.00、△RSCU≥0.08这2个条件[31]。
2 结果与分析
2.1 密码子碱基组成分析
普通核桃和深纹核桃的叶绿体基因组呈双链环状,为典型的四分体结构(图1、图2),普通核桃大单拷贝区(LSC)为89 872 bp,小单拷贝区(SSC)为18 423 bp,反向重复区包括IRA和IRB,均为26 036 bp;深纹核桃LSC为89 871 bp,SSC为18 412 bp,IRA为26 034 bp,IRB为26 034 bp。普通核桃的叶绿体基因组一共注释到136个基因,包括88个蛋白质编码基因,40个tRNA基因和8个rRNA基因,其中光合作用的相关基因包括6类[5个光系统Ⅰ基因、13个光系统Ⅱ基因、12个还原型辅酶Ⅰ(NADH)-脱氢酶基因、6个细胞色素b/f复合物基因、6个ATP合酶基因、1个RuBisCO亚基基因],自我复制的相关基因包括5类(15个小核糖体蛋白基因、11个大核糖体蛋白基因、4个RNA聚合酶基因、40个tRNA基因、8个rRNA基因),其他类型基因15个。深纹核桃的叶绿体基因组一共注释到131个基因,包括86个蛋白质编码基因,37个tRNA基因和8个rRNA基因,其中光合作用的相关基因包括6类[5个光系统Ⅰ基因、14个光系统Ⅱ基因、12个NADH-脱氢酶基因、6个细胞色素b/f复合物基因、6个腺嘌呤核苷三磷酸(ATP)合酶基因、1个RuBisCO亚基基因],自我复制的相关基因包括5类(15个小核糖体蛋白基因、10个大核糖体蛋白基因、4个RNA聚合酶基因、37个tRNA基因、8个rRNA基因),其他类型基因13个。
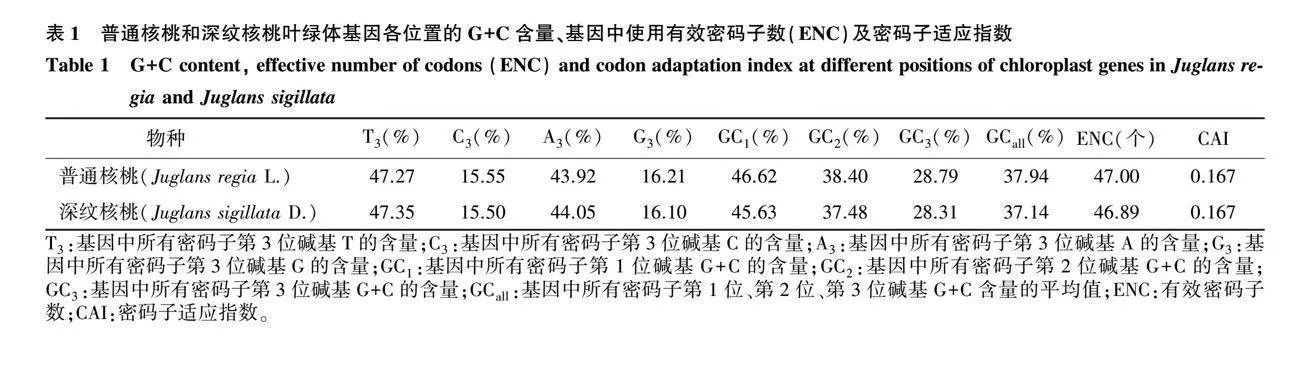
利用GeneiousPrime对2种核桃的叶绿体基因组序列进行筛选,删除长度过短或重复的CDS,选择典型基因序列,最终普通核桃和深纹核桃各获得53条CDS。进一步分析,普通核桃密码子第3位碱基上T、C、A、G的含量分别为47.27%、15.55%、43.92%、16.21%;深纹核桃密码子第3位碱基上T、C、A、G的含量分别为47.35%、15.50%、44.05%、16.10%,上述2种核桃中T3和A3高于G3和C3(表1)。
普通核桃的ENC范围为37.68~58.08个,深纹核桃的ENC范围为37.85~58.90个,进一步分析发现,共有74条(普通核桃37条、深纹核桃37条)叶绿体基因组序列的有效密码子数大于45.00个,表明2种核桃叶绿体密码子使用偏好性较弱,从表2可以看出,GC在不同位置、不同基因中未均匀分布。
对密码子各参数进行相关性分析,结果(表3)表明,在普通核桃和深纹核桃中,ENC与GC3极显著相关(P<0.01),与GC1、GC2相关性不显著,说明GC3对密码子偏好性存在较强影响;GC1、GC2与GC3相关性不显著,说明3个位置的碱基组成差异较大。
进一步对普通核桃和深纹核桃叶绿体基因组中各53条CDS进行分析,2种核桃RSCU大于1.00的密码子数量均为32个,普通核桃中以U结尾且RSCU>1的密码子有UUU、CUU、AUU、GUU、UCU、UAU、CAU、AAU、GAU、UGU,以A结尾且RSCU>1的密码子有AUA、GUA、UCA、CCA、ACA、GCA、UAA、CAA、AAA、GAA、CGA、AGA、GGA,以G结尾、RSCU>1的密码子有UUG、UCG、CCG、GCG、UAG、CGG、AGG、GGG,以C结尾且RSCU>1的密码子为UCC(表4)。深纹核桃中不同之处之一是以A结尾的密码子不含CGA,而含有UUA。由此可见,2种核桃以A和U结尾的密码子出现频率较高,为偏好密码子。
2.2 中性绘图分析
图3显示,普通核桃叶绿体基因组密码子的GC12、GC3的取值范围分别为0.333~0.543、0.210~0.365,深纹核桃叶绿体基因组密码子的GC12、GC3的取值范围分别为0.306~0.543、0.210~0.365,由此可见,2种核桃的叶绿体基因组密码子分布基本重合,说明普通核桃和深纹核桃中叶绿体基因组密码子的G+C含量接近。普通核桃GC12与GC3的回归系数为0.379 7,决定系数为0.016 9;深纹核桃GC12与GC3的回归系数为0.343 7,决定系数为0.028 2。2种核桃回归系数均不接近1.000 0,说明G+C在第1位、第2位、第3位这3个位置的碱基组成差异大,GC12与GC3相关性不显著。大部分基因位于对角线上方,综合相关性分析和中性绘图分析的结果,2种核桃叶绿体基因组密码子的使用偏好性主要受自然选择的影响。
2.3 ENC-plot绘图分析
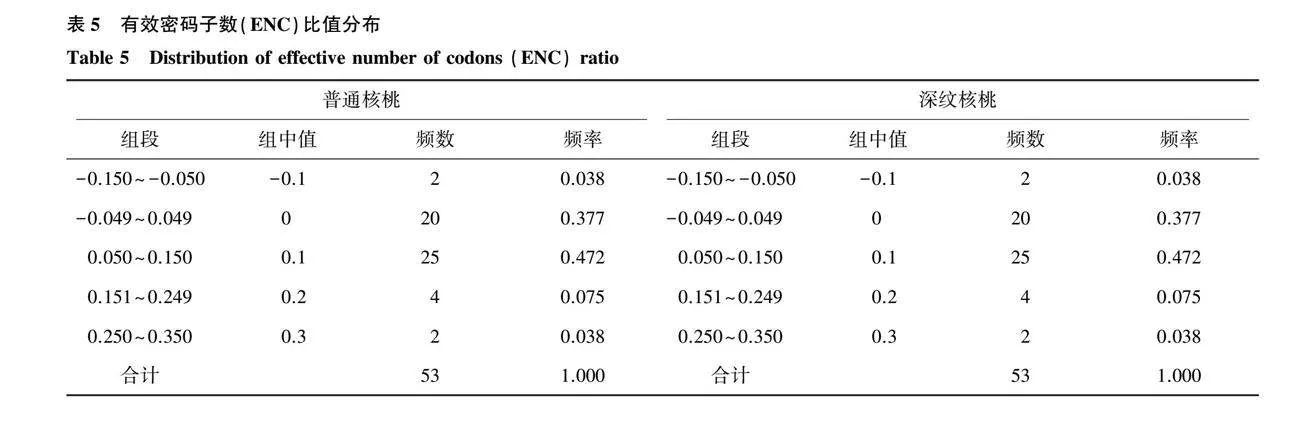
ENC-plot分析结果(图4)表明,少数基因位于标准曲线上,该部分基因的ENC实际观察值与ENC期望值差异较小;大部分基因远离标准曲线,其ENC实际观察值与ENC期望值相差较大。进一步分析基因与标准曲线的距离,并对ENC比值进行计算(表5),发现普通核桃和深纹核桃叶绿体密码子基因组在-0.049~0.049区间的基因频率为0.377,ENC实际观察值与ENC期望值差距较大。上述结果表明普通核桃和深纹核桃叶绿体密码子使用偏好性主要受到自然选择的影响。
2.4 PR2-plot分析
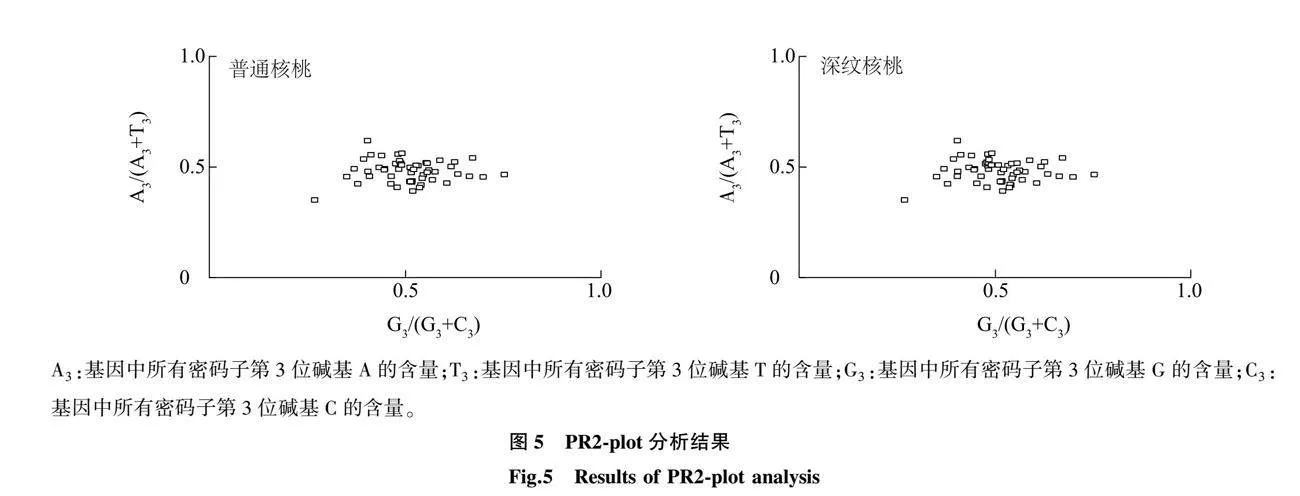
密码子偏好性仅受基因突变影响时,第3位碱基A/T、G/C具有相等的概率;受到自然选择影响时,A/T、G/C的概率不均等。对上述CDS进行碱基奇偶性分析,结果(图5)表明,2种核桃叶绿体基因在4个区域的基因位点呈现不均匀分布,在A3/(A3+T3)<0.5和G3/(G3+C3)>0.5区域分布得最多,表示密码子在第3位碱基的选择上具有偏好性,使用频率表现为T>A、G>C。上述结果说明,普通核桃和深纹核桃叶绿体基因组密码子偏好性受到自然选择的影响。
2.5 最优密码子的确定
对5个高表达基因petD、rps18、rps14、cemA、petB以及5个低表达基因atpF、ndhK、rpl2、ycf2、ycf3中的同义密码子进行计算,结果(表6)表明,普通核桃中有24个密码子(其中11个以U结尾,6个以A结尾,3个以G结尾,4个以C结尾)的△RSCU(高表达基因的RSCU-低表达基因的RSCU)≥0.08,深纹核桃中有26个密码子(其中11个以U结尾,8个以A结尾,3个以G结尾,4个以C结尾)△RSCU≥0.08。结合表4,普通核桃和深纹核桃中RSCU>1.00的密码子均为32个。同时满足RSCU>1.00和△RSCU≥0.08这2个条件的密码子为最优密码子,从普通核桃中筛选出UUU、AUU、UUG、GUU、CUU、UCC、UCG、CCG、GCA、UAU、CAA、AAA、GAA、UGU共14个最优密码子;从深纹核桃中筛选出UCC、UCA、UCG、CCG、UUU、GCA、UUA、UAU、UUG、CAA、CUU、AAA、AUU、GAA、AUA、UGU、GUU共17个最优密码子,上述共有23个最优密码子在第3个位置偏好以碱基A/U结尾。
3 讨论
密码子作为生物体内重要的遗传信息,在核糖核酸翻译为蛋白质的过程中发挥着重要作用。前人研究发现,在自然环境中,生物体中同义密码子的使用频率逐渐适应宿主的生长环境,并出现不能均衡使用的情况[32-33]。密码子使用偏好性受不同因素(基因序列碱基组成、自然选择等)影响[34],但一般认为自然选择是影响密码子使用偏好性的主要因素[35]。前人研究多将第3位碱基的G+C含量作为评价密码子偏好性的关键参数[36]。ENC能反映同义密码子非均衡使用的偏好情况,是最具参考价值的参数,ENC理论取值范围为20~61[37]。
普通核桃和深纹核桃的叶绿体基因组密码子中第1位、第2位和第3位碱基的组成都影响着叶绿体CDS的使用偏好。第3位碱基G+C含量总体小于第1位、第2位碱基,第3位碱基A、T含量均高于G、C含量,表明第3个位置上的碱基偏好使用A/T,这与南欧大戟(Euphorbia peplus L.)[33]、蒲桃属(Syzygium Gaertn.)[38]、杜鹃(Rhododendron simsii Planch.)[39]的密码子使用偏好性相同。碱基奇偶性分析结果表明,在第3个位置上碱基使用频率表现为T>A、G>C,说明2种核桃叶绿体基因组密码子使用偏好性受自然选择的影响,这与对木兰[40]、柿树[41]、蔷薇科果树[42]、紫花苜宿[43]等植物的分析结果相同;赵森等[44]分析秋茄,得出C大于G、T大于A的结果,说明秋茄叶绿体基因密码子使用模式受到自然选择的影响;竹节参叶绿体基因组密码子的偏好分析结果表明,基因突变是影响其密码子偏好形成的主要因素[45],这与本研究结果不一致,说明不同植物密码子使用偏好性受到的主要影响因素不尽相同。
4 结论
基于获得的普通核桃、深纹核桃叶绿体基因组序列,进行碱基组成、中性绘图分析、PR2-plot、RSCU值及ENC-plot分析比较,明确2种核桃拥有UUU、UUG、CUU、AUU、GUU、UCC、UCG、CCG、GCA、UAU、CAA、AAA、GAA、UGU共14个共同的最优密码子,多数以A或者U结尾,反映了相同分类单位、亲缘关系近的生物密码子偏好使用类似的碱基结尾[46-47]。基因突变和自然选择共同影响普通核桃、深纹核桃叶绿体基因组密码子的使用偏好性,但自然选择为主要影响因素。普通核桃中有14个最优密码子,深纹核桃中有17个最优密码子,且最优密码子更倾向于以A/U结尾,本研究得到2种核桃叶绿体基因组表达的重要信息,对之后进行核桃物种进化及种质资源的研究有着重要作用,同时也为预测核桃未知蛋白质的基因、基因工程表达载体的设计等提供了一定的数据支持。
参考文献:
[1] NOVOA E M, PAVON-ETERNOD M, PAN T, et al. A role for tRNA modifications in genome structure and codon usage[J]. Cell,2012,149(1):202-213.
[2] 赵春丽,彭丽云,王 晓,等. 苋菜AtGAI基因密码子偏好性与进化分析[J]. 中国农业大学学报,2019,24(12):10-22.
[3] 辛雅萱,黎若竹,李 鑫,等. 杧果叶绿体基因组密码子使用偏好性分析[J]. 中南林业科技大学学报,2021,41(9):148-156,165.
[4] 胡桂兵,张上隆,徐昌杰,等. 不同种类柑橘的密码子用法分析[J]. 华南农业大学学报,2006,27(1):13-16.
[5] HERSHBERG R, PETROV D A. Selection on codon bias[J]. Annual Review of Genetics,2008,42:287-299.
[6] 刘慧敏,乌云塔娜,杜红岩. 杜仲转录组基因密码子使用偏好性分析[J]. 北方园艺,2016(13):85-89.
[7] XU C, CAI X N, CHEN Q Z, et al. Factors affecting synonymous codon usage bias in chloroplast genome of Oncidium Gower Ramsey[J]. Evolutionary Bioinformatics,2011,7:271-278.
[8] 王 茹,罗永坚,刘相泉,等. 水杉叶绿体基因组同义密码子使用偏好性[J]. 湖北民族大学学报(自然科学版),2022,40(2):121-128.
[9] SINGH N D, DAVIS J C, PETROV D A. X-linked genes evolve higher codon bias in Drosophila and Caenorhabditis[J]. Genetics,2005,171(1):145-155.
[10]PARVATHY S T, UDAYASURIYAN V, BHADANA V. Codon usage bias[J]. Molecular Biology Reports, 2022,49(1):539-565.
[11]SHAW J, LICKEY E B, SCHILLING E E, et al. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms:the tortoise and the hare Ⅲ[J]. American Journal of Botany,2007,94(3):275-288.
[12]李西文,胡志刚,林小涵,等. 基于454FLX高通量技术的厚朴叶绿体全基因组测序及应用研究[J]. 药学学报,2012,47(1):124-130.
[13]冯 展,江 媛,郑 燕,等. 肉苁蓉属植物叶绿体基因组密码子偏好性分析[J]. 中草药,2023,54(5):1540-1550.
[14]谭淳月,刘 勇,赖章凤,等. 茶树氧甲基转移酶基因密码子偏好性分析[J]. 江西农业大学学报,2023,45(3):652-662.
[15]沈莲文,田金红,王玉昌,等. 2种玉兰属植物叶绿体基因组密码子偏好性分析[J]. 西南林业大学学报(自然科学),2023,43(2):44-53.
[16]原晓龙,刘 音,康洪梅,等. 蒜头果叶绿体基因组密码子偏好性分析[J]. 西南林业大学学报(自然科学),2021,41(3):15-22.
[17]夏 晞,彭劲谕,王大玮,等. 3种榕属叶绿体基因组密码子偏好性分析[J]. 西北林学院学报,2022,37(5):88-94.
[18]舒军霞,杨 林,周 涛,等. 刺柏属4种药用植物叶绿体基因组密码子偏好性分析[J]. 中草药,2022,53(23):7507-7515.
[19]孟 佳,方晓璞,史宣明,等. 我国核桃产业发展现状、问题与建议[J]. 中国油脂,2023,48(1):84-86,103.
[20]郭从善. 核桃及其加工与利用[J]. 粮油食品科技,1999(5):23-25.
[21]祝友朋,韩长志,熊 智. 核桃细菌性黑斑病菌分泌蛋白质的理化性质及特征分析[J]. 江苏农业学报,2019,35(2):295-301.
[22]谢 蕾,李贤忠,宁德鲁,等. 深纹核桃种质资源评价与挖掘研究进展[J]. 林业科技通讯,2021(11):26-31.
[23]覃 阳,全绍文,周 丽,等. 基于转录组测序的核桃(Juglans regia L.)SSR标记开发[J]. 分子植物育种,2019,17(20):6736-6742.
[24]余务刚,肖良俊,吴 涛,等. 不同类型深纹核桃的表型多样性研究[J]. 西部林业科学,2022,51(6):31-38,78.
[25]NOVEMBRE J A. Accounting for background nucleotide composition when measuring codon usage bias[J]. Molecular Biology and Evolution,2002,19(8):1390-1394.
[26]陆奇丰,骆文华,黄至欢. 两种梧桐叶绿体基因组密码子使用偏性分析[J]. 广西植物,2020,40(2):173-183.
[27]SUEOKA N. Directional mutation pressure and neutral molecular evolution[J]. Proceedings of the National Academy of Sciences of the United States of America,1988,85(8):2653-2657.
[28]YANG X, LUO X N, CAI X P. Analysis of codon usage pattern in Taenia saginata based on a transcriptome dataset[J]. Parasit Vectors,2014,7:527.
[29]SUEOKA N. Near homogeneity of PR2-bias fingerprints in the human genome and their implications in phylogenetic analyses[J]. Journal of Molecular Evolution,2001,53(4/5):469-476.
[30]唐晓芬,陈 莉,马玉韬. 密码子使用偏性量化方法研究综述[J]. 基因组学与应用生物学,2013,32(5):660-666.
[31]SHARP P M, LI W H. The codon adaptation index-a measure of directional synonymous codon usage bias,and its potential applications[J]. Nucleic Acids Research,1987,15(3):1281-1295.
[32]韩利红,代冬琴,赵明玉,等. 双孢蘑菇基因组密码子的偏好性分析[J]. 菌物学报,2021,40(3):603-615.
[33]余 涛,蒲 芬,管 芹,等. 南欧大戟叶绿体基因组密码子偏好性分析[J]. 江苏农业科学,2023,51(15):35-41.
[34]ROMERO H, ZAVALA A, MUSTO H. Codon usage in Chlamydia trachomatis is the result of strand-specific mutational biases and a complex pattern of selective forces[J]. Nucleic Acids Research,2000,28(10):2084-2090.
[35]吴学俊,梁湘兰,易子群,等. 湖北山楂叶绿体基因组密码子偏好性分析[J]. 分子植物育种,2023,21(8):2508-2516.
[36]冯德江,蔡华雅,刘 翔,等. 马铃薯X病毒外壳蛋白的表达水平与变偶密码子使用频率的相关性[J]. 微生物学报,2003,43(5):569-576.
[37]原晓龙,李云琴,张劲峰,等. 降香黄檀叶绿体基因组密码子偏好性分析[J]. 广西植物,2021,41(4):622-630.
[38]杨秀瑶,张梦洁,尹 拓,等. 蒲桃属叶绿体基因组特征及密码子偏好性分析[J]. 西南农业学报,2023,36(9):1869-1880.
[39]万露露,范敦锦,王中煊,等. 杜鹃叶绿体基因组特征及密码子偏好性分析[J]. 江苏农业科学,2023,51(13):54-59.
[40]季凯凯,宋希强,陈春国,等. 木兰科叶绿体基因组的密码子使用特征分析[J]. 中国农业科技导报,2020,22(11):52-62.
[41]傅建敏,索玉静,刘慧敏,等. 柿属植物叶绿体蛋白质编码基因密码子用法[J]. 经济林研究,2017,35(2):38-44.
[42]刘兴跃,何仲坚,邱毅敏. 四种蔷薇科果树叶绿体基因组密码子偏好性分析[J]. 分子植物育种,2022,20(16):5299-5308.
[43]喻 凤,韩 明. 紫花苜蓿叶绿体基因组密码子偏好性分析[J]. 广西植物,2021,41(12):2069-2076.
[44]赵 森,邓力华,陈 芬. 秋茄叶绿体基因组密码子使用偏好性分析[J]. 森林与环境学报,2020,40(5):534-541.
[45]梁 娥,齐敏杰,丁延庆,等. 竹节参转录组使用密码子偏好性分析[J]. 江苏农业科学,2019,47(2):59-63.
[46]李显煌,杨生超,辛雅萱,等. 灯盏花叶绿体基因组密码子偏好性分析[J]. 云南农业大学学报(自然科学),2021,36(3):384-392.
[47]王 婧,王天翼,王罗云,等. 沙枣叶绿体全基因组序列及其使用密码子偏性分析[J]. 西北植物学报,2019,39(9):1559-1572.
(责任编辑:王 妮)
