葡萄VvLBD35和VvLBD36的生物信息学与表达分析
2024-09-22李颖李高青高明刚许瑞瑞

摘要:LATERAL ORGAN BOUNDARIES DOMAIN(LBD)家族是含有高度保守的LOB结构域的一类植物特有转录因子,参与植物的生长代谢以及多种逆境胁迫应答等过程。本研究通过生物信息学方法对葡萄LBD基因家族中的VvLBD35和VvLBD36进行基因结构和进化关系分析,并利用qRT-PCR技术检测两个基因的组织表达模式以及在干旱、高盐、高温和低温条件下的诱导表达模式。结果显示,VvLBD35和VvLBD36均含有α-螺旋、无规则卷曲和β-转角结构,VvLBD36还含有较多的3-折叠结构;VvLBD35和VvLBD36与拟南芥、水稻、玉米、柑橘和番茄等植物中的LBD蛋白都有较高的同源性;VvLBD35和VvLBD36的启动子区含有多种胁迫、激素以及生长发育相关的应答元件:VvLBD35和VvLBD36在根、茎、叶中均有不同程度的表达,且均在叶中表达量最高,约是根和茎中的2-8倍:诱导表达模式分析表明,VvLBD35受高温诱导最为显著,下调至约为对照的6%,而VvLBD36受高盐和高温的诱导最为显著,分别上调至对照的5.67倍和14.29倍。本研究为进一步深入揭示VvLBD35和VvLBD36的生物学功能奠定了实验基础。
关键词:葡萄;LBD家族:序列分析;表达分析;非生物胁迫
中图分类号:S663.1:Q786 文献标识号:A 文章编号:1001-4942(2024) 08-0023-07
LBD转录因子家族,又称为AS2(asymmetricleaves 2-like)/LOB基因家族,因其编码蛋白中均含有一段LOB (lateral organ boundaries)结构域而得名。LOB结构域一般含有3种保守的基序,即类锌指结构域、Gly-Ala-Ser(GAS)结构域以及类亮氨酸拉链结构域。LBD转录因子家族根据LOB结构域的完整性可分为Ⅰ类和Ⅱ类。Ⅰ类包含3种完整的结构域,而且GAS结构域与类亮氨酸拉链结构域之间能够通过形成“螺旋卷曲”结构,介导蛋白质之间的相互作用。Ⅱ类具有一个残缺的亮氨酸拉链结构域,但Ⅱ类基因家族较Ⅰ类相对保守。LBD基因家族中Ⅰ类成员较多。研究显示,LBD蛋白间一般以形成异源二聚体的形式发挥生物学作用。
目前,已在多种植物中鉴定到LBD基因家族成员,在拟南芥中鉴定到43个、大麦中24个、番茄中46个、玉米中44个、毛果杨中57个LBD基因。LBD转录因子参与植物的生长代谢以及多种逆境胁迫应答等过程。拟南芥中鉴定到AtLBD16、AtLBD29在生长素响应因子ARF7介导的侧根发育以及温度依赖性愈伤组织的形成中均发挥着关键作用:AtLBD15通过参与ABA信号转导途径调控植物的干旱胁迫应答:AtLBD27参与拟南芥器官的发生过程;AtLBD18、AtLBD30、AtLBD36等在拟南芥导管分化、细胞壁形成和花发育中均可发挥重要的作用:AtLBD37、AtLBD39调控拟南芥花青素的形成和氮响应过程。PtLBD1参与调控杨树次生生长过程。小立碗藓受甘露醇诱导后大多数PpLBDs表达量均上调,可能会增加植株的耐旱性。菊花CmLBD1的表达受生长素调节并参与不定根原基的形成过程。农作物中也鉴定到一些LBDs广泛地参与到对非生物胁迫的应答以及植物生长发育调控过程。比如,玉米ZmLBD2和ZmLBD5均参与干旱胁迫响应的调控过程,ZmLBD19调控植物器官发生的过程:马铃薯StLBD2 -6和StLBD3-5的表达受干旱胁迫诱导,可能参与干旱胁迫应答:水稻Ⅱ类LBD蛋白OsLBD37和Os-LBD38介导抽穗期的生长调控:番茄SlLBD40介导茉莉酸信号转导负调控干旱胁迫应答:小麦TaLBD16 - 4D参与植株结构和抽穗期的调控:大豆GmLBD12经干旱和多种激素等胁迫处理后被显著诱导。然而,与拟南芥和农作物相比,经济作物葡萄LBD基因家族的研究却相对较少,与逆境胁迫的关系更是鲜有报道。
葡萄作为一种具有较高营养价值的经济作物,种植历史悠久,在全世界范围内被广泛种植。然而,全球气候变暖、工业污染、干旱、土壤盐碱化等非生物胁迫的加剧严重影响着葡萄的生长过程。因此,研究葡萄胁迫应答相关基因、挖掘耐胁迫基因资源,为后续有针对性地进行胁迫应答相关基因的改造以培育具有多种胁迫抗性的新品种有重要的理论价值和现实意义。
前期研究通过生物信息学和同源性分析鉴定到葡萄中存在40个LBD基因(VvLBDl -VvLBD40),本研究通过生物信息学方法分析葡萄LBD基因家族成员VvLBD35和VvLBD36的基因结构特征和进化关系,利用实时荧光定量PCR(qRT-PCR)检测两者在葡萄根、茎、叶中的组织表达模式以及在干旱、高盐、高温和低温条件下的诱导表达模式,为进一步揭示VvLBD基因家族应答逆境胁迫的生物学作用提供理论基础。
1 材料与方法
1.1 试验材料和处理
葡萄品种为‘贵妃’,于2021年9月在日光温室中按照品种最适条件进行常规管理。葡萄种子首先置于4℃条件下层积,2d后移入25℃、16h光照/8 h黑暗的培养室中培养。分别收集培养4周后葡萄植株的根、茎、叶组织,经液氮速冻后移入-80℃超低温冰箱保存。
诱导表达试验:采用干旱(200 mmol/L甘露醇)、高盐(300 mmol/L NaCl)、高温(42℃)和低温(4℃)分别处理生长4周的葡萄植株6h;以正常条件下的葡萄植株作为对照。胁迫处理后收集葡萄叶片并立即用液氮冷冻保存。
1.2 VvLBD35和VvLBD36蛋白的亚细胞定位及三级结构预测
从Phytozome数据库(http://www.phytozome.org/)下载葡萄VvLBD35和VvLBD36的核苷酸和氨基酸序列,通过查阅LBD转录因子保守基序的研究结果,依据其保守基序的序列特征,分析绘制VvLBD35和VvLB’D36的基因序列图:使用WoLFPSORT在线网站(https://www.genscript.com/wolf-psort.html)进行VvLBD35和VvLBD36的亚细胞定位预测;利用SWISS-MODEL(http://swissmod-el.expasy. org)在线预测葡萄VvLBD35和Vv-LBD36蛋白的三级结构。
1.3 VvLBD35和VvLBD36的序列比对和系统进化树构建
采用DNAMAN生物学软件对葡萄VvLBD35与拟南芥AtLBD27、水稻Os0890402100、玉米2mOOOOleb042180、柑橘CcLBD6、番茄SILBD27进行氨基酸保守域序列比对,对VvLBD36与拟南芥AtLBD38、水稻OsLBD37、玉米2mOOOOleb326200、柑橘CICLE_v10026374mg、番茄Solyc029092550进行氨基酸保守域序列比对。利用ClustalX软件对葡萄、拟南芥、水稻、玉米、柑橘、番茄6个物种的蛋白序列进行分析比对。利用MEGA7.0构建系统进化树。
1.4 VvLBD35和VvLBD36的顺式作用元件分析
利用PlantCARE(http://bioinformatics. psb.ugent. be/webtools/plantcare/html/)在线预测葡萄VvLBD35和VvLBD36启动子区(约2 000 bp)存在的顺式作用元件,最后利用TBtools软件进行可视化绘图。
1.5 葡萄总RNA提取与qRT-PCR检测
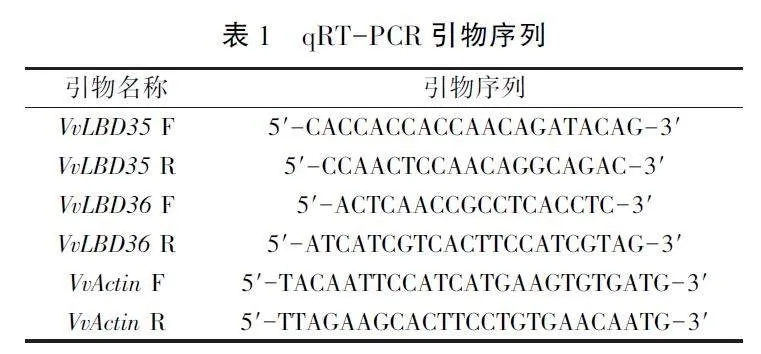
利用RN03 RNA提取试剂盒(北京Aidlab公司)提取葡萄组织总RNA,然后反转录合成cD-NA。qRT-PCR检测采用Bio-Rad CFX96系统,使用Takara的SYBR Green PCR Master Mix试剂盒。VvActin作为内参基因,测定VvLBD35和Vv-LBD36在葡萄不同组织以及不同胁迫处理下叶片中的表达模式,重复3次。引物信息见表1。
1.6 数据处理与分析
利用2 -AAC‘法计算VvLBD35和VvLBD36基因的相对表达量。利用SPSS 20.0软件进行方差分析,采用最小显著差数法(LSD)进行多重比较。
2 结果与分析
2.1 VvLBD35和VvLBD36蛋白亚细胞定位及三级结构分析
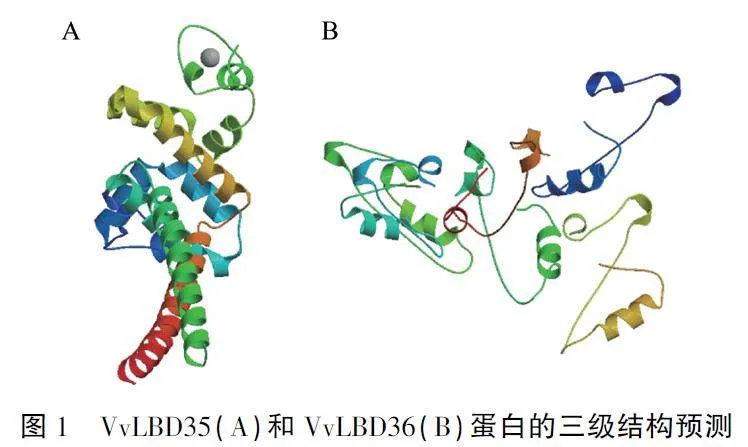
VvLBD35和VvLBD36分别编码301、222个氨基酸,对其编码蛋白进行结构分析。结果显示,VvLBD35具有典型的C-block基序CAACK-YQRRKCSSEC,结构模式符合CX2CX6CX3C,属于LBD基因家族的Ⅰ类:而VvLBD36则不具有以上基序特征,含有一个残缺的亮氨酸拉链结构,表明其属于Ⅱ类。亚细胞定位预测结果显示,Vv-LBD35和VvLBD36可能均定位于细胞核。蛋白质三级结构分析结果(图1)显示,VvLBD35包含多个α螺旋,无规则卷曲和β-转角较少,未发现β-折叠;VvLBD36包含少量的α螺旋,较多的β-折叠、β-转角和无规则卷曲结构。
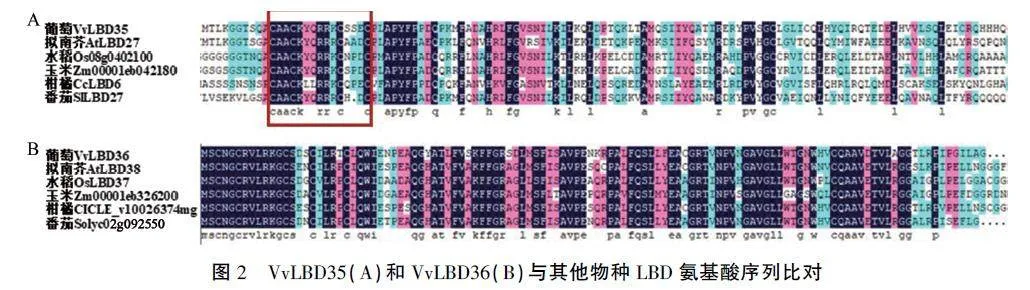
2.2 VvLBD35和VvLBD36的氨基酸序列比对和系统进化分析
如图2所示,通过对VvLBD35和VvLBD36蛋白与其他5个物种LBD家族成员分别进行氨基酸多序列比对,VvLBD35与拟南芥AtLBD27、水稻Os08g0402100、玉米2m0000leb042180、柑橘CcLBD6、番茄SILBD27均包含保守LOB结构域中的C基序,该序列的相似性高达85%,全部序列的相似性为33%(图2A);VvLBD36与拟南芥AtLBD38、水稻OsLBD37、玉米2mOOOOleb326200、柑橘CICLE_v10026374mg、番茄Solyc02g092550的序列相似性达到61%(图2B)。表明6种LBD家族成员的N端区域高度保守,C端区域具有特异性,为可变C端。图2 VvLBD35(A)和VvLBD36(B)与其他物种LBD氨基酸序列比对
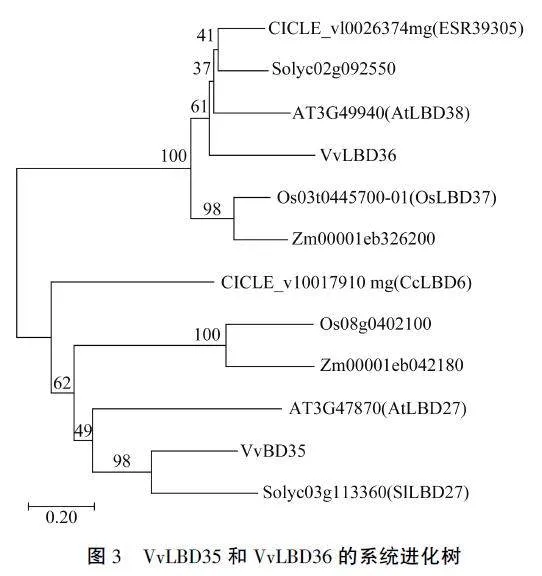
为了进一步了解VvLBD35和VvLBD36转录因子的进化关系,利用MEGA7.0软件对12种LBD蛋白进行系统进化分析。结果(图3)显示,VvLBD35转录因子与番茄SILBD27、拟南芥AtLBD27同源性较高,系统进化关系较近,其次是玉米2m00001eb042180、水稻Os08g0402100以及柑橘CcLBD6; VvLBD36与拟南芥AtLBD38、番茄Solyc02g092550以及柑橘CICLE_v10026374mg同源性较高,系统进化关系较近,其次是水稻Os-LBD37和玉米2m00001eb326200。
2.3 VvLBD35和VvLBD36启动子顺式作用元件分析
为了解VvLBD35和VvLBD36的潜在功能和调控机制,利用PlantCARE在线软件分别提取葡萄VvLBD35和VvLBD36约2 000 bp的启动子序列,并进行顺式作用元件分析。如图4所示,Vv-LBD35启动子上分布有脱落酸作用元件(ABRE)、茉莉酸甲酯作用元件(CGTCA -motif)、生长素反应元件(TGA - element)、光响应元件(TCCC-motif)、低温反应元件(LTR)以及干旱胁迫反应元件(MBS)等;VvLBD36启动子区域则含有赤霉素(P-box)、水杨酸(TCA-element)、氧胁迫(GC -motif)和茉莉酸甲酯(CGTCA - motif和TGACG-motif)响应等相关的顺式作用元件;同时,这两个基因的启动子中还含有与生长发育相关的元件,如胚乳发育元件(GCN4_motif)和分生组织表达元件(CAT-box)等。表明VvLBD35和VvLBD36可能在葡萄的非生物胁迫应答和生长发育方面均发挥着重要作用。
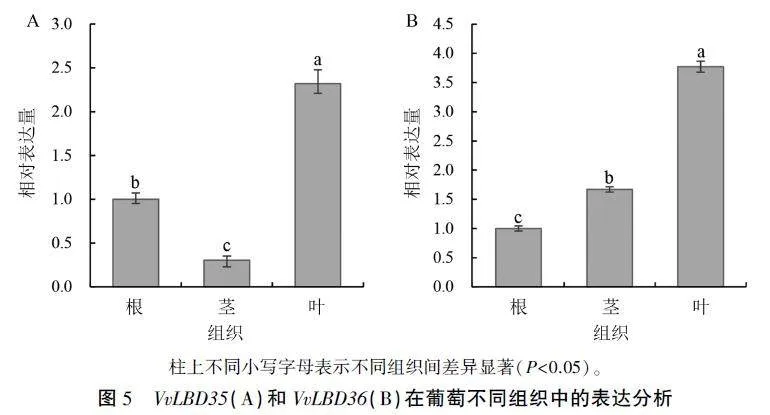
2.4 VvLBD35和VvLBD36的组织表达模式分析
为分析VvLBD35和VvLBD36在葡萄不同组织中的表达情况,以生长4周的葡萄不同组织的cDNA为模板,利用qRT-PCR技术分别检测Vv-LBD35和VvLBD36在葡萄根、茎和叶中的表达量。结果(图5)显示,VvLBD35和VvLBD36在根、茎和叶中均有表达,且均在叶中表达量最高,约是根和茎中的2-8倍。可见,VvLBD35和VvLBD36在葡萄不同组织中表达程度不同,这可能与其在不同组织中的表达调控有关。
2.5 VvLBD35和VvLBD36的诱导表达模式分析
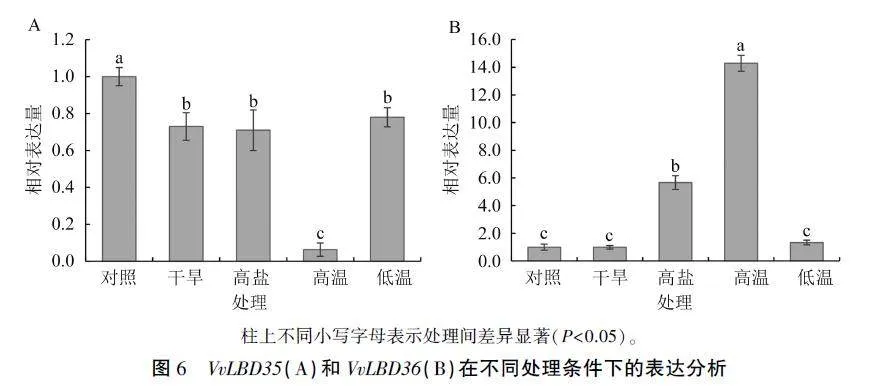
VvLBD35和VvLBD36在不同诱导处理下的表达模式分析结果显示,与对照相比,各诱导条件F VvLBD35的表达量均显著降低,以高温条件下降低最显著,下调至约为对照的6%(图6A);Vv-LBD36受高盐和高温条件诱导表达水平显著升高,分别为对照的5.67倍和14.29倍,而干旱和低温处理几乎不影响VvLBD36的表达(图6B)。
3 讨论与结论
转录因子作为转录调控最后一环的关键因子,是近年来植物逆境胁迫分子机制研究中的一个热点。葡萄在栽培种植过程中极易遭受逆境胁迫,为更加有效地控制非生物胁迫对葡萄的危害,提高经济效益,葡萄的抗逆性尤其是耐低温、耐高温、干旱抗性以及响应盐害的能力亟待提高。为了能更好地了解葡萄转录因子LBD蛋白的结构特征和表达调控特点,本研究对葡萄Vv-LBD35和VvLBD36进行了蛋白特征的预测,分析了两个基因在葡萄主要组织中的内源表达规律以及不同胁迫条件下的诱导表达模式,为深入揭示VvLBD35和VvLBD36响应逆境胁迫的调控机制和生物学功能奠定了基础。
LBD基因已经在许多植物中被鉴定且大部分成员都属于I类。毛果杨57个LBD基因中I类占79%:水稻基因组中鉴定到35个LBD基因,其中I类成员占比高达86%:小立碗藓基因组中鉴定到31个LBD转录因子,属于I类的成员占比为77%。前人由于研究策略、查询序列、数据库选择等差异在葡萄基因组中分别鉴定到了40、50、30个LBD家族成员,而且经过基因分析发现大部分成员也归属于I类家族。本研究通过对葡萄VvLBD35和VvLBD36基因进行蛋白特征分析得知,VvLBD35属于I类,具有CX,CX6CX3C的结构模式:而VvLBD36则不具有以上基序特征,只含有1个残缺亮氨酸拉链结构,证明Vv-LBD36转录因子属于Ⅱ类亚家族。
截至目前,LBD转录因子在多个物种响应非生物胁迫中发挥的作用已被验证。拟南芥AtLBD15、番茄SILBD40、马铃薯StLBD1 -5以及小立碗藓PpLBDs等参与了植物的干旱胁迫应答;大豆GmLBD12受干旱、低温等多种因素的显著诱导:16个BnLBDs能够提高苎麻植株的耐热性:辣椒中I类LBD基因在响应高温胁迫时表达量明显低于Ⅱ类LBD基因。
通过对葡萄VvLBD35和VvLBD36启动子区的顺式作用元件分析发现,二者启动子区域均含有与非生物胁迫应答相关的元件。诱导表达模式分析显示,高温条件下VvLBD35的表达显著下调,同样VvLBD36受高温胁迫的诱导表达最为显著,表明,VvLBD35和VvLBD36可能在植物响应高温胁迫中发挥重要作用。另外,诱导表达模式分析表明葡萄VvLBD36也受到高盐胁迫的诱导。本研究结果为进一步揭示葡萄LBD转录因子在高温等非生物胁迫应答中的作用和抗性机制奠定了理论基础。
基金项目:山东省自然科学基金面上项目(ZR2022MC064);作物生物学国家重点实验室开放课题(2021KF06);潍坊学院博士科研基 金项目(2023BS19)
