小麦大、小粒材料的细胞学鉴定及其粒重相关基因表达研究
2024-09-22徐晓艺汪晓璐韩冉马朋涛徐文竞刘成


摘要:籽粒大小是影响小麦产量的重要农艺性状,开展控制籽粒大小相关基因的筛选鉴定和表达研究对小麦高产育种具有借鉴意义。本研究利用染色体计数和荧光原位杂交(FISH)的方法对15份大粒材料(千粒重高于50 g)和15份小粒材料(千粒重低于21 g)进行鉴定,结果显示,京5044、京5085、藏1783、黑72-0484、uis鉴010共5份大粒材料的染色体条数为28,为四倍体小麦:其余材料染色体条数均为42,为六倍体小麦。选取六倍体的大粒和小粒材料各3份作为试验材料,用水稻和拟南芥的64个粒重相关基因在小麦基因组中筛选同源基因,对其进行qRT-PCR分析,选出6个在大、小粒材料中表达量存在显著差异的基因进行对比分析。结果表明,基因RiGSK2、RiGW8和RiGGC2在大粒材料HR2015-1-1- 2-9中表达量较高,RiGS3和RiD11在小粒材料红秃头3中表达量较高,ArTTG2在3份小粒材料中表达量均较高,推测这6个基因可能是控制小麦粒重的关键基因,在后续研究中将进一步验证它们对小麦产量的贡献。
关键词:小麦:籽粒大小:荧光原位杂交(FISH):粒重相关基因:表达分析
中图分类号:S512.1:Q786 文献标识号:A 文章编号:1001-4942(2024) 08-0010-07
种子大小是植物最重要的产量性状之一,因此,开展控制籽粒大小的基因的相关研究,对培育高产小麦具有重要意义。目前已从小麦中鉴定出一些籽粒大小相关基因,主要包括负调控粒宽和千粒重的TaGW2基因及其同源基因,负调控种子大小的泛素激活蛋白酶基因TaDA1,编码具有重复类似Kelch - like结构的磷酸激酶基因TaGL3,调节细胞分裂素稳态的细胞分裂素氧化酶基因TaCKX,参与淀粉合成的关键基因TaACP-S1,控制粒宽和粒长的小麦蔗糖合成酶基因TaSUS1,正调控籽粒大小和重量的编码丝氨酸羧肽酶的基因TaGS5,与粒重、粒宽、粒长相关的谷氨酰胺合成酶基因TaGS1,以及其他一些参与小麦籽粒大小形成的基因如TaC-YP78A3、TaGS3等。然而小麦基因组复杂庞大,仍存在大量重要的控制大小籽粒形成的基因未被挖掘出来,因此,继续深入挖掘籽粒大小与重量相关基因对于小麦品种改良意义重大。
目前控制种子大小的信号途径在拟南芥和水稻中研究较多,已经发现了如泛素—蛋白酶体途径、G蛋白信号途径、丝裂原活化蛋白激酶信号途径、植物激素和转录调控因子途径等多种相关信号途径。研究发现,丝裂原活化蛋白激酶信号途径中,拟南芥MKK4/5在MAPK6的上游控制胚形成:转录调控因子途径中,拟南芥TTG2编码一个WRKY家族转录因子,导致ttg2突变体被皮中的细胞长度减小:泛素一蛋白酶体途径中,水稻GW2编码一个环型E3泛素连接酶,通过加速籽粒灌浆,增加粒宽、粒重和产量;G蛋白信号途径中,水稻RGB1 -DEP1/GGC2二聚体和分离的RGA1可能通过促进穗壳中的细胞增殖激活未知的下游效应因子来调节籽粒的大小。但以上基因是否参与控制小麦大小粒发育的信号通路仍不清楚。
本研究以15份大粒和15份小粒小麦材料为基础,通过染色体计数和FISH核型分析选出含42条染色体、千粒重最高的3份大粒材料和最低的3份小粒材料,利用64个控制水稻和拟南芥大小籽粒发育的基因,通过同源比对的方法从小麦基因组中鉴定出同源基因,并用qRT-PCR法分析其在6份大、小粒材料中的表达差异,筛选出控制小麦籽粒大小的重要基因,为后续验证这些基因对小麦产量的贡献并用于品种改良提供依据。
1 材料与方法
1.1 供试材料
千粒重超过50 9的15份大粒小麦材料:DA-LI-1、DALI-2、京5044、京5085、藏1783、黑72-0484由山东省农业科学院作物研究所(以下简称作物所)张荣志副研究员提供;DASUIDALI、LJ15鉴010和章丘大穗由作物所刘建军研究员提供:HR2015-1-1-2-9、咸阳超大穗101、咸阳超大穗362、咸阳超大穗901、咸阳超大穗965由作物所韩冉博士提供:大颖壳由四川农业大学甯顺宗教授提供。
千粒重低于21 9的15份小粒材料:红秃头3、红穗、白穗白、小白麦、小白穗、腰珠红、芦麦、改麦、亮麦、小秃头、红芒麦、红半芒、红秃头1、齐头子和红秃头2为地方品种,由作物所樊庆琦研究员提供。
1.2 植物材料培养及样品处理
将30份供试小麦材料的种子放于铺有湿润滤纸的培养皿中,置于22-24℃恒温光照培养箱中培养约24 h。待种子萌发后,将培养皿转到4℃冰箱处理24 h。将处理过的种子转入22 - 24℃恒温光照箱中培养24-35 h,待根尖长至1.0-1.5 cm时采集根尖,用冰水混合物处理24 h后用卡诺固定液I(无水乙醇:冰醋酸=3:1)固定3-7 d,放人4℃冰箱备用。
将筛选出的3份大粒材料HR2015-I-I-2-9、大颖壳、咸阳超大穗101和3份小粒品种白穗白、红穗和红秃头3的种子种在10 cm×10 cm的花盆中,每个品种种5盆,每盆6株,在光照培养室培养(24℃,湿度60%,14 h光照/10 h黑暗);孕穗期取幼嫩小穗,液氮处理后于-80℃冰箱保存备用。每份材料选3株长势一致的单株作为3个生物学重复。
1.3 千粒重的调查统计
2021年9月在山东省农业科学院试验田(36°2'N,117°5'E)种植15份大粒材料和15份小粒材料,2022年6月成熟期采收并调查千粒重。
1.4 染色体计数与FISH核型分析
将备用根尖的分生组织切下,放人含2%果胶酶R-IO(日本Yakult公司)和2%纤维素酶Y-23(日本Yakult公司)的10 μL混合酶液中,37℃水浴50 min酶解;酶解后,用70%乙醇冲洗2次,然后用解剖针捣碎,2 000 r·min-1离心10 s,倒掉乙醇,加入100%乙酸20 μL,涡旋混匀;取7μL根尖分生组织细胞悬浮液,滴到载玻片上,停留约5 min待乙酸完全挥发后镜检,镜检合格的染色体制片置于-20℃冰柜中备用。
取浓度约为10μmol·L-1的双色FISH探针(Oligo-pSc119.2-1和Oligo-pTa535-1)各0.2 μL加入到8 μL 2x柠檬酸钠(SSC)溶液(pH=7)中,涡旋混匀配成杂交液。将杂交液滴至上述制备好的玻片上,盖上盖玻片,置于湿润的塑料盒中80℃变性5 min,然后将塑料盒置于37℃杂交箱中杂交Sh。取出染色体制片,放于装有2xSSC溶液的洗脱缸中,洗掉盖玻片和杂交液,然后再用ddH,O冲洗一遍,晾干,避光滴加一滴抗褪色剂,盖上盖玻片,用OLYMPUS BX53荧光显微镜(奥林巴斯,日本)镜检,DP70CCD镜头下拍照。
1.5 RNA提取及qRT-PCR分析
将采集的3份大粒材料和3份小粒材料的幼嫩小穗分别利用miRcute多糖多酚植物miRNA提取分离试剂盒(天根,北京)提取RNA,利用Nanodrop 2000/2000C分光光度计(Thermo,美国)测定RNA浓度。用PrimeScript RT reagent Kitwith gDNA Eraser(TaKaRa,大连)将RNA反转录成cDNA,用于qRT-PCR分析。
选取已报道的34个控制水稻籽粒大小发育和30个控制拟南芥籽粒大小发育的基因,在NC-BI网站(https://www.ncbi. nlm. nih. gov)搜索相应的小麦同源基因,根据其保守序列区段,通过Primer3Plus软件设计qRT-PCR引物,对其在3份大粒材料和3份小粒材料幼嫩小穗中的表达量进行qRT-PCR测定,发现有6个基因的表达量在大粒和小粒材料间存在显著差异,对这6个基因在大、小粒材料中的表达情况进行对比分析。这6个基因的引物序列见表1。PCR反应体系为20 μL,包括:SYBR@ Green PCR Master Mix 10μL,稀释的cDNA 2 μL,0.1 μmol·L-1正、反向引物各0.4 μL,用RNase free water补齐至20 μL。PCR反应程序:95℃预变性5 min;95℃变性10s,60℃延伸30 s,40个循环。设置3次技术重复,用2-△△Ct法计算基因相对表达量。利用SPSSStatistics 21.0软件进行数据统计分析。
2 结果与分析
2.1 供试小麦材料的细胞学鉴定
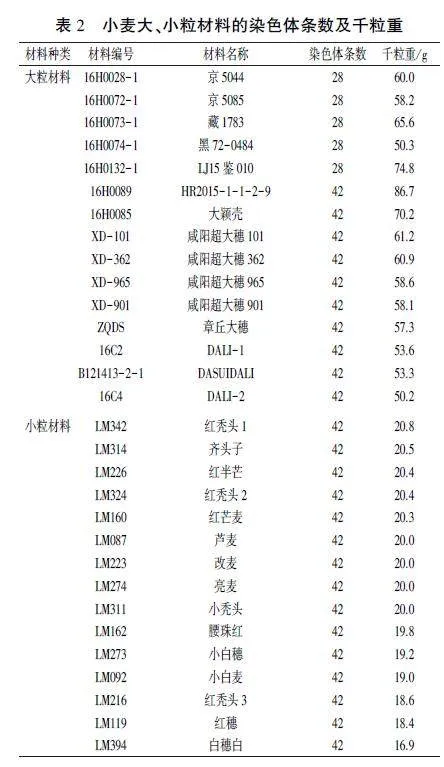
利用染色体计数和FISH法对15份大粒材料和15份小粒材料进行细胞学分析,结果(表2)显示,京5044、京5085、藏1783、黑72-0484、LJ15鉴010共5份大粒材料的染色体为28条,是四倍体小麦:其余大粒材料及所有小粒材料的染色体均为42条,是六倍体小麦。FISH鉴定结果以四倍体小麦黑72-0484和六倍体小麦HR2015 -1-1-2-9为例展示,见图1。
2.2 用于分析粒重相关基因表达的小麦材料筛选
2022年6月收获供试小麦材料,统计千粒重,结果见表2,大粒材料的千粒重在50.2 - 86.7 9范围内,小粒材料的千粒重在16.9 - 20.8 9范围内。从染色体条数为42的材料中,选千粒重高的前3位大粒材料以及千粒重低的前3位小粒材料,分别为HR2015-1-1-2-9、大颖壳和咸阳超大穗101以及白穗白、红穗和红秃头3,用于后续粒重相关基因分析。
2.3 粒重相关基因在小麦大、小粒材料中的表达分析
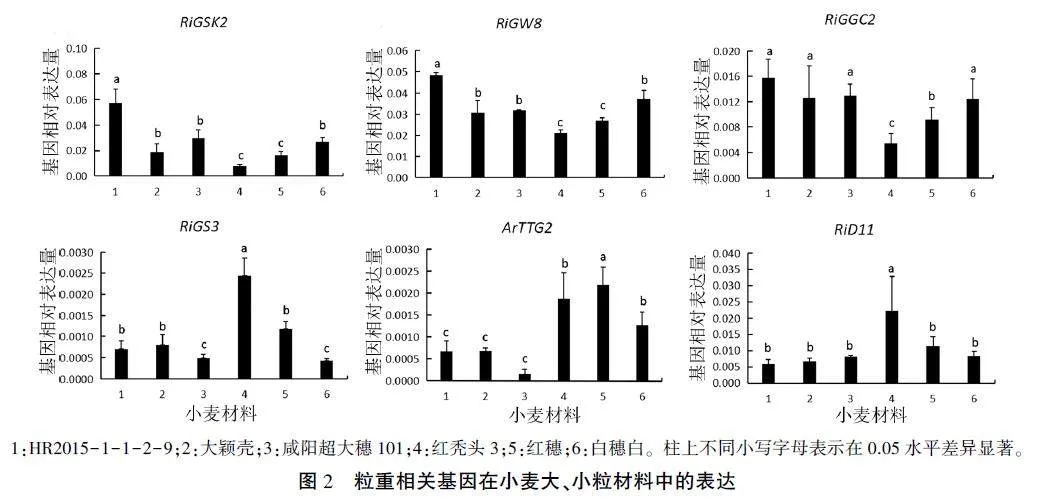
利用64个控制水稻和拟南芥大小籽粒发育的基因,通过同源比对的方法从小麦基因组中找出同源基因,对其在3份大粒材料和3份小粒材料中的表达情况进行qRT-PCR分析,其中有6个基因在大、小粒材料中的表达有显著性差异,如图2显示。RiGSK2和RiGW8均在HR2015 -1-1-2-9中的表达量最高,且均显著高于其他材料,分别为大粒材料大颖壳、咸阳超大穗101的3.0、1.9倍和1.6、1.6倍,分别为小粒材料红秃头3、红穗、白穗白的8.1,3.6、2.2倍和2.3、1.8、1.3倍;RiG-GC2也是在HR2015-1-1-2-9中的表达量最高,显著高于在红秃头3和红穗中的表达量,约是两者的2.9倍和1.3倍,但与其他材料差异不显著。上述3个基因均在红秃头3中的表达量最低,在红穗中的表达量较低。RiGS3在红秃头3中的表达量最高,显著高于其他材料,约是小粒材料红穗和白穗白的2.0倍和5.6倍,依次是大粒材料HR2015-1-1-2-9、大颖壳、咸阳超大穗101的3.4、3.0、4.8倍;其在白穗白和咸阳超大穗101中的表达量显著低于其他材料。ArTTG2、RiD11在小粒材料中的表达量高于大粒材料,且ArTTG2在3份小粒材料中均达到差异显著水平,而RiD11仅在红秃头3中达到差异显著水平:ArTTG2在红穗中的表达量最高,分别是红秃头3和白穗白的1.2倍和1.7倍,依次是大粒材料HR2015 -1-1-2-9、大颖壳、咸阳超大穗101的3.1、3.1、14.0倍;RiD11则是在红秃头3中表达量最高,分别是红穗和白穗白的2.0倍和2.7倍,依次是大粒材料HR2015-1- 1- 2-9、大颖壳、咸阳超大穗101的3.3、3.7、2.8倍。
3 讨论与结论
3.1 控制籽粒大小的小麦同源基因在大粒材料中的作用
小麦产量由单位面积穗数、穗粒数和千粒重三要素决定,其中千粒重是影响产量的关键因素。挖掘小麦籽粒大小关键基因对于提高粒重进而提高产量意义重大。如:Su等根据水稻中OsGV2基因调控粒重,同源克隆了小麦中与粒重相关的基因TaGW2。Ma等克隆了TaGS5 -3A基因,发现TaGS5-3A -T比TaGS5-3A -G基因型有更高的酶活:过表达该基因后,小麦籽粒增大,千粒重提高。Li等发现过表达CKX2导致拟南芥iku2种子增大。可见,挖掘并过表达控制大粒发育的基因可提高小麦产量。
本研究利用水稻和拟南芥中调控籽粒发育的基因进行同源比对,筛选到6个小麦同源基因,通过其在小麦大、小粒材料中的表达分析发现,RiG-SK2、RiGW8和RiGGC2可能为控制小麦大粒形成的基因,其在3份大粒材料中的表达量显著高于小粒材料红秃头3:RiGSK2、RiGW8在HR2015-1-1-2-9中的表达量显著高于其他两份大粒材料和3份小粒材料。这可为小麦高产育种提供重要的基因资源,今后可将其用于小麦的高产育种。
3.2 控制籽粒大小的小麦同源基因在小粒材料中的作用
Hong等利用RNA干扰(RNAi)技术,降低了调控小粒形成的TaGV2A、TaGW2B、TaGV2D基因的转录水平,使得小麦粒宽、粒重增加。Wang等[28]利用CRISPR/Cas924产生OsOTUB1敲除突变体,使得水稻穗粒数和千粒重增加,从而提高了籽粒产量。Zhao等验证发现转基因水稻OsGSK2的RNAi系(OsGSK2-Gi-2)使得水稻籽粒长度增加了22%。可见,挖掘调控小粒形成基因,通过基因编辑干扰其表达可以提高作物产量。本研究发现,RiGS3、ArTTG2、RiD11基因可能与小麦小粒形成有关。其中ArTTG2在3份小粒材料中的表达量均显著高于3份大粒材料:RiGS3、RiD11在红秃头3中的表达量显著高于3份大粒材料,与其他两份小粒材料红穗及白穗白之间也存在显著差异。因此可通过基因编辑技术将小粒形成基因RiGS3、ArTTG2和RiD11用于小麦高产育种,实现小麦增产。
综上,本研究利用控制水稻和拟南芥大小籽粒发育的基因,从小麦基因组中鉴定出同源基因,并利用qRT-PCR法分析了各籽粒大小相关基因在不同大粒和小粒材料中的表达差异,初步筛选到可能控制大粒发育的基因RiGSK2、RiGW8和RiGGC2以及控制小粒发育的基因RiGS3、ArT-TG2、RiD11,为进一步验证其功能及调控机制奠定了基础,同时可为小麦高产分子育种提供重要的基因资源。
基金项目:山东省自然科学基金项目(ZR202IQC198,ZR2020MC098);山东省重点研发计划项目(2022LZG002)
