核桃晚霜危害特性及影响机制研究进展
2024-06-28张绥林李洋李琰张赟齐齐建勋侯智霞
张绥林 李洋 李琰 张赟齐 齐建勋 侯智霞
摘要:早春晚霜冻害的发生给我国核桃生产带来严峻的考验,严重影响核桃生长发育、产量及收益。为明确我国核桃晚霜危害特性及影响机制,综述了核桃受晚霜危害程度的影响因素及其在响应低温胁迫时生理生化指标变化和分子机制。分析发现,关于生理变化相关的研究集中在细胞膜系统(相对电导率)、抗氧化系统(丙二醛、超氧化物歧化酶、过氧化物酶及过氧化氢酶)和渗透调节物质(可溶性糖、可溶性蛋白质及脯氨酸)3个方面,JrPAL、JrJAZ、JrGRAS、JrICE1、JrCBF/JrDREB1、JrCOR、JrWRKY、JrDHN、JrGST 和JrPUB 等基因的表达量受到低温胁迫的诱导。对未来的研究方向进行了展望,为进一步挖掘核桃应对晚霜危害的响应机制及采取有效的生产措施提供了科学依据。
关键词:核桃;晚霜危害;低温胁迫;影响因素;机理
doi:10.13304/j.nykjdb.2022.0952
中图分类号:S664.1 文献标志码:A 文章编号:10080864(2024)04001809
核桃(Juglans regia L.)是胡桃科(Juglandaceae)胡桃属(Juglans L.)落叶乔木,也是世界四大干果之一。核桃生长适宜的年平均温度为9~16 ℃,极端低温不低于-25 ℃ ,极端高温不超过35~38 ℃[1]。我国北方地区核桃的花期一般在4—5月[2]。‘薄壳山核桃的生物学零度为10 ℃,从萌芽到果实成熟需要有效积温3 300 ℃以上,雌雄花从花芽萌动至终花期期间≥10 ℃有效积温在458.0~579.7 ℃有利于‘薄壳山核桃开花[2]。晚霜又称“倒春寒”,当早春温度较高且持续时间较长,外界环境温度突然≤0 ℃时,晚霜就会发生[3]。晚霜危害多发生在核桃的花期至幼果发育早期,通常情况下,外界温度高于0 ℃的晚霜冻害会导致核桃幼叶发黑,但不会对已经展叶的幼茎造成伤害;然而,外界温度低于0 ℃的晚霜冻害会导致核桃嫩芽、花器官、幼茎等受冻部位发黑、萎蔫,甚至脱落[3]。近年来,我国北方核桃产区春季晚霜频发,其发生时间往往与核桃花期和幼果期相遇,导致相关器官受害,严重时直接影响核桃的产量和经济效益[45]。核桃新梢耐受低温临界值为-2~-4 ℃,花、幼果耐受低温临界值为-1 ℃[6]。此外,核桃受晚霜危害程度与低温持续时间有关,低温持续时间越长,核桃受害程度越重[3]。不同地区核桃受晚霜危害的程度通常用该年核桃损失的产量进行评估,一般认为产量损失<10% 为轻度冻害,≥10% 且<30% 为中度冻害,≥30% 且<60% 为重度冻害,≥60% 为危重冻害[7]。目前,晚霜危害对于核桃芽、花等器官的影响以及核桃对造成低温伤害的响应机制尚不明确,这将严重影响抗(避)晚霜核桃种质科学选育的效率,也不利于核桃应对晚霜危害有效措施的研发。因此,本研究从我国核桃产区晚霜危害的影响因素出发,通过对细胞膜系统、抗氧化系统、渗透调节物质等生理生化特性的变化进行分析,结合低温环境下相关基因的表达,对核桃响应晚霜危害的分子机制进行了综述,有助于了解核桃应对晚霜的响应机制,指导核桃产业的生产实践活动,实现增产增收,为我国核桃产业带来一定的经济效益。
1 核桃受晚霜危害程度的影响因素
1.1 核桃产区气候
晚霜对核桃产量的危害程度受到地理位置的影响。云南、新疆、四川及陕西是我国核桃的主产区,我国北方核桃产区温度普遍低于南方,发生晚霜的频率也高于南方核桃产区,尤其是新疆喀什、陕西宜君县、山西晋中、甘肃成县等地晚霜发生频繁,对核桃造成的晚霜危害较严重[3]。多年来,我国各地的晚霜多在4月中旬发生,新疆喀什市通常在4月12日左右[8],2006年晚霜导致喀什大约117 hm2核桃受害[9],严重影响核桃产业的经济收益。陕西省铜川市晚霜通常在4月上中旬发生,2020年4月10—12日晚霜造成宜君县核桃受灾面积达194.466 7 hm2,经济损失高达225.06 万元[4]。陕西省宜君县2018年发生晚霜冻害的强度大于2020 年,2018 年4 月极端最低温度为-1.8 ℃,≤0 ℃低温持续时间为5 h,而2020年没有晚霜冻害记录[4]。陕西省蓝田县灞源在4 月上、中、下旬发生霜冻的次数分别为15、7、4次,而灞源正处于山区,可能是因为山区的高海拔和低气温导致其霜冻频率较高[5]。上述说明,陕西省不同产区核桃受晚霜危害程度与当地低温及其持续时间关系密切。
山西省早实核桃通常于4月初萌动,花芽于4月上中旬萌动。2015年4月13日,山西省蒲县、汾阳和黎城的最低温度分别为-4、0和0 ℃,早实核桃产量损失率分别为60%、10%和20%,说明核桃受害程度与所处环境温度呈反比[6]。同时,晚实核桃产量损失率分别为70%、8%和15%,可能是因为-4 ℃的低温环境导致晚实核桃顶花芽受冻,当年无法开花结果,而早实核桃受冻后产生的二芽、三芽可以正常萌发结果[6]。2020年清明节前后的晚霜冻害使山西省晋中市太谷地区核桃受冻率高达30%,2018年4月上旬的晚霜冻害导致太谷地区核桃受冻面积达95%以上[10]。2018年4月上旬,甘肃成县晚霜导致核桃受到不同程度的冻害,其中二郎镇、王磨镇、小川镇、沙扒镇达到危重冻害程度,陈院镇、纸坊镇、索池乡、苏园乡受到严重冻害,城关镇、抛沙镇受到重度冻害[11]。然而,目前关于晚霜危害发生情况的统计并不完全,对于核桃主产区具体种植地区核桃产量受晚霜冻害影响的统计仍然存在较大缺口。因此,后续研究中详细统计每个核桃种植地区的晚霜危害特点以及产量、经济损失等具体情况对于明确我国核桃受晚霜冻害程度具有重要意义。此外,基于不同核桃产区的气候特点,深入探索各产区核桃能够承受晚霜危害的临界值,对于挖掘核桃响应晚霜危害的机制也至关重要。
1.2 核桃品种
晚霜危害通常导致核桃受害器官或整株发育延缓、叶片变黄枯萎、分蘖数减少、结果率降低[12],甚至导致花苞、幼果和新梢枯死。不同核桃品种物候期各异,导致其受晚霜冻害影响程度不一。山西晋南地区晚霜基本在3月上旬至3月底发生,而‘鲁光‘辽宁1号和‘礼品2号在3月18—19日进入萌动期,恰逢晚霜发生时间,上述核桃品种花芽因受冻而变黑、干枯、脱落,影响当年核桃产量[13]。‘清香‘上宋6号的雌花盛花期在5月,‘汾核系列‘礼品2号的雌花盛花期在4月底,而山西吕梁核桃产区的晚霜通常于4月上中旬发生,上述核桃品种可以有效避开晚霜冻害的影响[14]。美国核桃品种‘强特勒‘土莱尔和‘美国黑核桃在甘肃成县萌芽较晚,恰能避开4月上旬的晚霜冻害[15]。当核桃的物候期避开晚霜发生日期时,其受晚霜冻害影响较小,也称为避晚霜品种,可以为抗(避)晚霜核桃品种的选育提供参考。选育抗(避)晚霜品种是解决核桃受晚霜冻害影响最直接的途径。因此,可以基于对不同品种抗性和核桃物候期的观察,并结合杂交育种或无融合生殖的手段,选育可以有效抗(避)晚霜危害的优质子代或新品种。
2 核桃受低温胁迫的生理生化机制
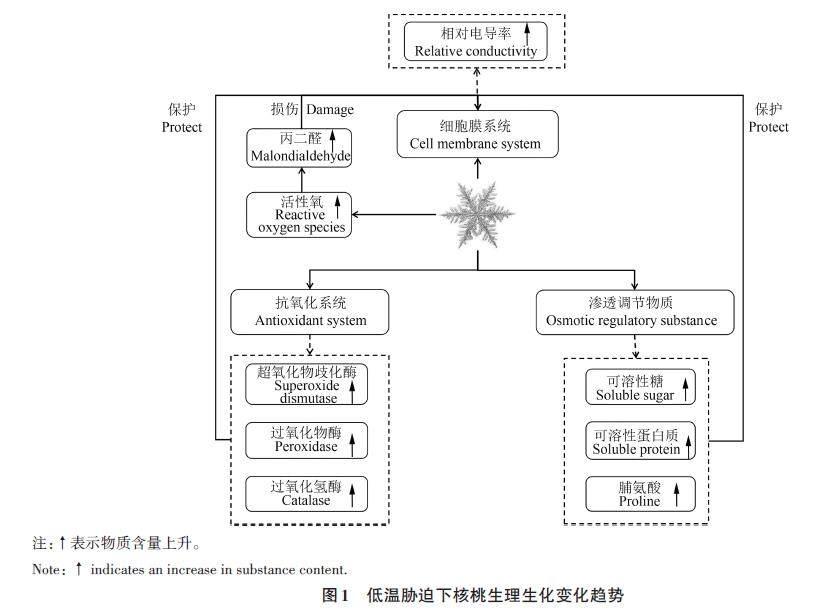
目前,大多研究均从抗寒性的角度评估核桃对低温胁迫的耐受能力[1617]。解析低温胁迫对核桃生理生化特性的影响往往从细胞膜系统、抗氧化系统及渗透调节物质3个方面开展(图1)[10]。
2.1 细胞膜系统
通常情况下,核桃新梢耐受低温临界温度为-2~-4 ℃,叶片、花、幼果耐受低温临界温度为-1 ℃[6]。当环境温度低于-1 ℃时,核桃花和幼果便会受冻而干枯、脱落;低于-4 ℃时,核桃新梢便会受冻而变黑、干枯。与此同时,细胞膜受到伤害后透性会增大。因此,相对电导率(relativeconductivity, REC)的变化可以反映细胞膜受害程度,进一步说明核桃受晚霜危害程度。当生长季核桃受到4 ℃低温胁迫时,叶片REC变化不大;而在-4 ℃低温胁迫时,REC显著升高[1819]。当新疆核桃‘扎343‘温185‘新新2号和‘露仁1年生枝条受到≤0 ℃低温胁迫时,其REC显著升高[16]。综上可知,当环境温度在核桃耐受低温临界值内时,REC保持相对稳定,低于临界值时,REC呈升高趋势。因此,当早春晚霜危害发生时,可以将REC作为判定核桃受晚霜危害程度的指标,用来评估核桃细胞膜结构和功能受损伤的程度。
2.2 抗氧化系统
当核桃受到≤0 ℃ 低温胁迫时,活性氧(reactive oxygen species, ROS)生成速率增加,核桃1 年生枝条中丙二醛(malondialdehyde, MDA)大量积累(图1),造成膜脂过氧化,导致ROS产生与清除之间的动态平衡遭到破坏,从而对核桃造成伤害[20]。当核桃叶片受到≤0 ℃低温胁迫时,其MDA含量随着温度的降低呈上升趋势[21]。综上可知,核桃受到≤0 ℃低温胁迫时,MDA含量增加表明细胞膜受到了严重损害。抗寒性越强的核桃品种受到≤0 ℃低温胁迫时,其体内MDA含量降低。例如,‘鲁果12 号枝条MDA 含量显著低于‘香玲[22],‘绿岭叶片中MDA含量显著低于其他核桃品种[21],表明‘鲁果12号和‘绿岭受害程度更低。
植物体内常见的抗氧化系统以抗氧化酶为主,主要包括超氧化物歧化酶(superoxidedismutase, SOD)、过氧化氢酶(catalase, CAT)和过氧化物酶(peroxidase, POD)[20]。SOD、POD和CAT是植物酶促防御体系中重要的保护酶,能防止膜脂过氧化。研究证明,SOD、POD和CAT在低温胁迫环境中具有特殊的变化规律。目前,SOD、POD、CAT在核桃应对≤0 ℃低温胁迫中的响应已经受到关注[17-20]。当‘香玲叶片和子房受到≤ 0 ℃低温胁迫时,随着温度的降低(0~-5 ℃),SOD及POD活性先升高后降低,叶片SOD及POD活性在-4 ℃达到峰值,子房SOD 及POD 活性在-3 ℃达到峰值[21]。1 ℃低温胁迫12 h时,‘哈特雷‘晋龙1号和‘晋龙2号叶片SOD和POD活性升高;随着低温胁迫时间的继续延长,SOD和POD活性呈下降趋势[23]。‘香玲和‘鲁果12号受到≤0 ℃低温胁迫后,1年生枝条中SOD活性呈“升-降-升-降”的趋势,CAT活性呈先上升后下降的趋势[22]。当温度降至一定程度(耐受低温临界温度)前,核桃体内SOD、POD 和CAT 活性在逐渐升高(图1)。而当温度继续下降时,SOD、POD和CAT活性开始下降。同时,≤0 ℃低温持续时间达到一定程度时,SOD、POD和CAT活性升高;超过该时间,SOD、POD 和CAT活性便会下降。≤0 ℃低温胁迫初期,SOD、POD 和CAT 活性的升高有利于核桃清除低温导致的自由基的积累,保护核桃细胞膜系统,进一步提升核桃的抗寒能力。而温度的继续下降及低温持续时间的延长打破了核桃体内的代谢平衡,SOD、POD 和CAT 活性的下降导致核桃清除自由基的能力显著下降,导致核桃受到低温冻害的影响。因此,在生产实践中,将核桃生长环境温度控制在一定范围(高于耐受低温临界温度)也是一种预防晚霜危害的有效措施。这些生理生化特性变化趋势为准确衡量核桃受晚霜危害程度提供了参考依据,SOD、POD和CAT活性与核桃受晚霜危害的关系及其分子调控机制值得进一步研究与挖掘。
2.3 渗透调节物质
目前,关于核桃受到≤0 ℃低温胁迫的研究涉及的渗透调节物质有可溶性糖(soluble sugar,SS)、可溶性蛋白(soluble protein, SP)和脯氨酸(proline, Pro),研究证明,SS、SP和Pro含量的变化可以用来鉴定核桃抵御低温胁迫的能力[17,20]。不同核桃品种受到≤0 ℃低温胁迫时,其体内SS、SP和Pro含量存在一定的差异[19]。例如,≤0 ℃低温胁迫下,避晚霜品种‘清香1年生枝条SS和SP含量显著低于不避晚霜品种‘辽宁1号[20]。抗晚霜品种‘温185 1年生枝条SS和SP含量显著高于其他品种,而Pro含量显著低于其他品种[24]。综上可知,较高的SS含量有利于清除核桃细胞内自由水含量,较高的Pro含量利于通过提高细胞渗透压而增加保水能力,较高的SP含量有利于提高核桃抵御≤ 0 ℃低温胁迫的能力(图1)。不同核桃品种渗透调节物质含量之间的差异性导致其抵御低温胁迫的能力存在差异,这对于后续选育抗晚霜核桃新品种具有重要的指导意义。
3 核桃受低温胁迫的分子机理
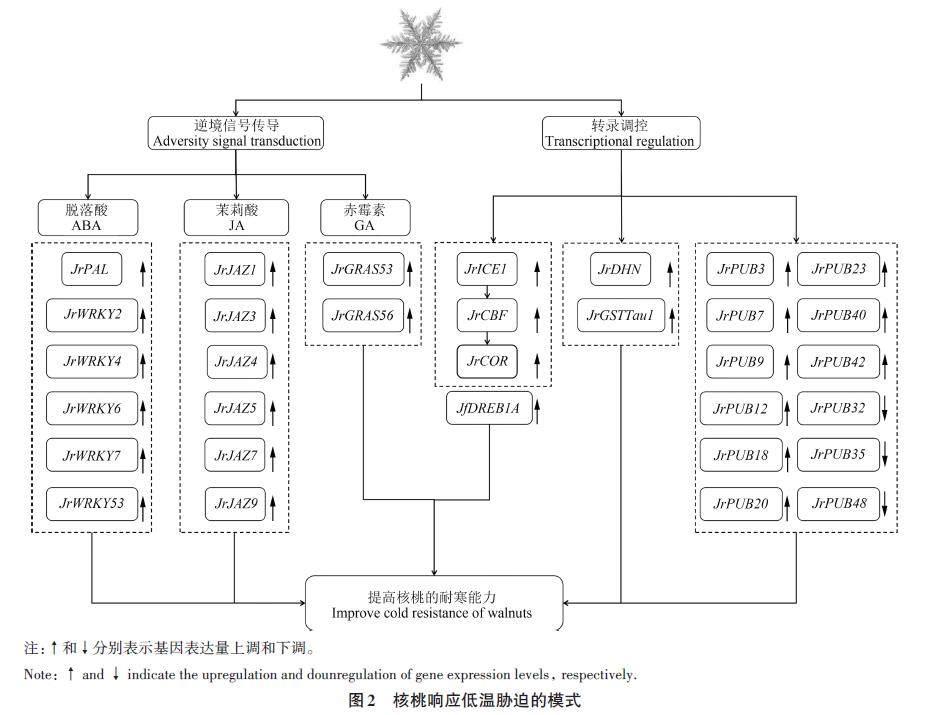
随着分子生物学技术水平的不断提高,核桃等经济林木响应低温胁迫分子机制的研究主要集中在低温保护物质和调节蛋白因子调控2 个方面[25]。在低温胁迫下,诱导响应低温胁迫基因的表达主要通过逆境信号传导、转录及转录后调节水平进行(图2)。
3.1 逆境信号传导
脱落酸(abscisic acid, ABA)是一种低温信号传导分子,ABA信号途径是植物处于低温胁迫等逆境反应的核心[26]。WRKY是一类植物特有的抗逆相关转录调控因子,已经被证明在植物响应低温等逆境胁迫中发挥重要作用[27]。5 ℃低温胁迫下,根系中JrWRKY2 的表达量先上升后下降,处理9 h后达到峰值,而JrWRKY7 在根系中的表达量呈“升-降-升”的趋势,处理2 h后达到峰值[26]。上述结果说明,JrWRKY 参与核桃受到低温胁迫的早期响应。外源ABA 使过表达JrWRKY2 和JrWRKY7 的核桃叶片中SOD 和POD 酶活性均显著高于对照、MDA含量显著低于对照。此外,5 ℃低温胁迫下,瞬时转化JrWRKY2 和JrWRKY7 的核桃叶片Evans蓝染色程度最浅,对照组叶片Evans蓝染色程度最深[26]。外源ABA 使核桃种子JrWRKY6和JrWRKY53表达量显著上升[27]。JrWRKY可能通过依赖于ABA的信号传导途径参与核桃对低温胁迫的响应,并提高其抗旱能力。研究表明,对4 ℃核桃苗进行外源ABA 处理后,其叶片JrPAL 表达水平上调,同时低温也可以诱导JrPAL(pyrabactin resistance 1-like)的表达[28],推测ABA可能通过调控JrPAL 提高核桃对低温环境的抵御能力。
JAZ(jasmonate ZIM-domain)蛋白是茉莉酸(jasmonic acid, JA)信号传导过程中重要的负调控蛋白,在调控植物生长和防御的JA信号传导中发挥着重要的作用[29]。0.5 ℃低温处理核桃苗48 h后,JrJAZ1、JrJAZ3、JrJAZ4、JrJAZ5、JrJAZ7 和JrJAZ9 在‘辽宁8号和‘清香叶片中的表达量均显著升高[29],即JrJAZ 参与核桃低温胁迫的响应。赤霉素(gibberellin, GA)合成代谢GRAS家族是存在于植物体内的转录调控因子,GRAS在赤霉素信号传导过程中发挥着重要作用,对低温胁迫做出响应[30]。对核桃GA信号传导途径响应低温胁迫的研究表明,4 ℃低温胁迫核桃苗48 h后,JrGRAS53 和JrGRAS56 在不耐低温核桃品种‘清香中下调,但在耐低温核桃品种‘辽宁8号中差异不显著[31]。对早实核桃‘中林1号和晚实核桃‘清香进行外源GA3处理,低质量浓度(50和100 mg·L-1)GA3处理有利于降低核桃受低温冻害的程度,而高质量浓度(200和300 mg·L-1)GA3处理增加核桃受低温冻害程度[32]。总之,GA 可能通过上调JrGRAS 而提高核桃抵御低温胁迫的能力。
3.2 转录调控
ICE (inducer of CBF expression) 属于bHLH(basic helix-loop-helix)转录因子,在模式植物拟南芥中,ICE1-CBF (C-repeat binding transcriptionfactor)-COR (cold-related)低温信号途径已经比较清晰[33],为核桃等木本植物响应低温胁迫机理的研究奠定了基础。当植物响应低温胁迫时,ICE1转录因子被激活,从而使其下游基因CBF 和COR上调表达,提高植物的抗寒能力[34]。‘清香嫁接苗受4 ℃低温胁迫48 h后,JrICE1 在其叶片中的表达量显著升高,表明JrICE1 参与核桃低温胁迫的响应[35]。随着4 ℃低温胁迫时间的延长,JrCOR 在‘香玲花芽中的表达量先上升后下降,并于处理4 h时达到峰值,说明JrCOR 主要在核桃响应低温胁迫的早期阶段发挥作用[36]。虽然目前已经研究了ICE1 和COR 基因家族在核桃受到4 ℃低温胁迫时的表达模式,但是ICE1、CBF 和COR 在核桃响应低温胁迫过程中的调控机制仍不清晰。
CBF/DREB1 (C-repeat binding factor/ dehydrationresponsive element binding protein 1)是植物体内低温信号传导途径中研究最广泛的转录因子,CBF/DREB1由低温诱导产生,有利于增强植物在逆境中的抗性[37]。核桃幼苗受4 ℃ 低温胁迫后,JfDREB1A 在根系、茎端和叶片中的表达量立即上调。此外,JfDREB1A 在拟南芥中的过表达通过维持细胞膜的稳定性、降低相对电导率和提高SOD、POD 和CAT活性来提高拟南芥的存活率。无性系核桃组培苗受到4 ℃低温胁迫后,叶片中JrCBF表达量随着低温胁迫时间的延长呈“先上升后下降”的趋势,在8 h时达到峰值,并在24 h时降至对照组的24 倍,进一步说明JrCBF 能够被低温诱导[38]。然而,在2和4 ℃低温胁迫下,‘阳光和‘云新这2个品种的核桃叶片中CBF 表达量没有显著差异[39],与徐丽等[38]的研究存在差异,可能是因为‘阳光和‘云新这2个核桃品种对低温胁迫的反应并不敏感。综上,CBF/DREB1 参与了核桃响应低温胁迫的生物学过程,但其分子机制尚不明确。
此外,DHN(dehydrins)、GSTTaul (glutathioneS-transferase)和PUB(U-box)等也在核桃受低温胁迫时发挥作用。4 ℃低温胁迫处理核桃幼苗,叶片中JrDHN 的表达量增加,并在处理4 h后达到峰值[40]。6 ℃低温胁迫下,JrGSTTau1 在核桃叶片中的表达量显著高于温度在6 ℃以上的叶片[41]。4 ℃低温胁迫处理核桃苗48 h后,叶片中JrPUB3(U-box)、JrPUB7、JrPUB9、JrPUB12、JrPUB18、JrPUB20、JrPUB23、JrPUB40 和JrPUB42 的表达量显著上调,而JrPUB32、JrPUB35 和JrPUB48 的表达量显著下调[42]。上述结果说明JrDHN、JrGST和JrPUBs 在核桃响应低温胁迫的过程中发挥着重要作用。
综上所述,一定程度的低温胁迫通过诱导低温胁迫响应相关基因的表达,进而提高核桃抵御低温胁迫的能力;ICE1、CBF/DREB1、COR、DHN、GST 和PUB 等在低温胁迫中发挥着重要作用,以上结果不仅有助于更好地从分子水平理解核桃对早春晚霜危害的响应,更有利于继续开展核桃抗晚霜机制的研究。然而,目前ABA、JA及GA调控核桃响应低温胁迫的作用机制尚不清楚,WRKY、JAZ及GRAS在核桃响应晚霜危害时的调控网络仍需进一步解析,在分子水平上对核桃抗晚霜以及相关基因家族提升核桃耐受低温胁迫的信号通路方面的研究也相对欠缺。因此,后续可以从ABA、JA 及GA 信号传导途径出发,基于WRKY、JAZ、GRAS、ICE1-CBF-COR、DHN、GST 和PUB 等基因关联研究核桃响应晚霜危害的具体分子调控机制。
4 展望
我国核桃主产区集中在云南、新疆、四川、陕西等地,大面积土地的优势有利于核桃产业增产增收,而近年来频发的晚霜冻害给我国核桃的生产带来了严峻的考验。目前,生产上已采取一系列的措施减轻核桃的晚霜危害并逐渐完善,通过选育抗(避)晚霜品种[9]、核桃受害后立即剪去所有受冻新梢的物理防治方法以及喷施赤霉素等激素推迟核桃花芽萌发的化学防治方法[33]降低核桃受害率,提升核桃产量和品质。虽然核桃受到低温胁迫时生理生化指标及其响应低温胁迫的分子机制已经取得一定的研究进展,而响应低温胁迫的相关基因的克隆及其在核桃植株中的功能鉴定等方面还处于起步阶段。挖掘核桃抗晚霜的分子响应机制对选育和改良抗晚霜核桃种质资源具有重要的指导意义。目前,使核桃有效避开晚霜冻害的措施主要集中在培育避晚霜冻害的新品种上。关于核桃抗晚霜研究可以从以下方面深入:①挖掘ABA、JA、GA等信号通路在核桃抗晚霜中的分子机制及调控网络;② 探索ICE、CBF/DREB1、COR 和WRKY 等响应低温胁迫基因参与核桃抗晚霜的调控机制;③通过对不同品种核桃物候期的观察,结合杂交育种技术,选育抗(避)晚霜核桃新品种;④基于随低温胁迫相关基因的克隆,通过构建核桃的遗传转化体系,结合基因编辑技术,培育抗(避)晚霜核桃新品种。
参 考 文 献
[1] 郗荣庭. 果树栽培学总论[M]. 北京:中国农业出版社,1997:303.
[2] 蒋瑶, 魏海林, 高昌虎, 等. 湖南低山丘陵区薄壳山核桃的开花物候期观测及品种配置[J]. 南京林业大学学报(自然科学版), 2021, 45(1): 53-62.
JIANG Y, WEI H L, GAO C H, et al .. Observation on floweringphenology and variety combination of Carya illinoinensis in lowmountains and hills of Hunan province [J]. J. Nanjing For.Univ. (Nat. Sci.), 2021, 45(1): 53-62.
[3] 王一峰, 陈耀年, 王瀚, 等. 我国核桃晚霜冻害研究进展[J].农业与技术, 2020, 40(11): 49-51.
[4] 杨亚利, 郑合清, 张淑敏, 等. 2018和2020年铜川核桃晚霜冻害分析[J]. 陕西气象, 2022(3): 64-68.
[5] 乐呼, 雷晓英, 袁晓霞, 等. 蓝田县核桃春季霜冻特征及风险区划[J]. 现代农业科技, 2022(5): 163-165,169.
[6] 武静, 贺奇, 史敏华, 等. 山西省核桃晚霜危害调查及防御措施研究[J]. 山西林业科技, 2016, 45(3): 28-30.
WU J, HE Q, SHI M H, et al .. Study on the damage andpreventive measures of late frost to Juglans regia in Shanxiprovince [J]. Shanxi For. Sci. Technol., 2016, 45(3): 28-30.
[7] 樊江斌, 张金龙, 许新平, 等. 陕西2013年核桃晚霜冻害发生情况调查与分析[J]. 西北林学院学报, 2014, 29(5):120-124.
PAN J B, ZHANG J L, XU X P, et al .. An investigation onfrozen damage of walnut trees in Shaanxi province [J]. J.Northwest For. Univ., 2014, 29(5): 120-124.
[8] 王荣梅, 张晓琴, 刘姣, 等. 新疆喀什地区近50 a来霜冻变化特征及其对农业的影响[J]. 干旱气象, 2013, 31(2):309-312.
WANG R M, ZHANG X Q, LIU J, et al .. Frost variation and itsinfluence on agricultural in Kashi of Xinjiang in recent 50years [J]. J. Arid Meteorol., 2013, 31(2): 309-312.
[9] 刘杜玲, 张博勇, 彭少兵, 等. 早实核桃物候期观察与避晚霜品种的筛选[J]. 北方园艺, 2011(24): 14-17.
LIU D L, ZHANG B Y, PENG S B, et al .. Observation onphenological period and varieties choice in avoiding the latefrost damage of early-fruiting walnut culivars [J]. NorthernHortic., 2011(24): 14-17.
[10] 武彦霞, 田鑫, 李夏媛, 等. 山西省核桃晚霜冻害及预防补救措施[J]. 果树资源学报, 2021, 2(1): 51-52.
WU Y X, TIAN X, LI X Y, et al .. Late frost damage of walnutand preventive and remedial measures in Shanxi province [J].J. Fruit Resour., 2021, 2(1): 51-52.
[11] 王一峰, 赵淑玲, 庞世伟, 等. 2018年成县核桃晚霜冻害调查与分析[J]. 陕西林业科技, 2018, 46(3): 32-35.
WANG Y F, ZHAO S L, PANG S W, et al .. Investigation onfreezing damage to walnut trees in Cheng county [J]. ShaanxiFor. Sci. Technol., 2018, 46(3): 32-35.
[12] YADAV S K. Cold stress tolerance mechanisms in plants. areview [J]. Agron. Sustain. Dev., 2010, 30(3): 515-527.
[13] 常月梅, 李永琛, 高俊仙. 核桃资源的抗晚霜性能评价[J].果树资源学报, 2022, 3(5): 15-19.
CHANG Y M, LI Y C, GAO J X. Evaluation of late frostresistance of walnut genetic resources [J]. J. Fruit Resour.,2022, 3(5): 15-19.
[14] 冀中锐, 张树振, 史根生, 等. 核桃物候期观察与避晚霜品种的筛选[J]. 落叶果树, 2021, 53(1): 13-16.
JI Z R, ZHANG S Z, SHI G S, et al .. Observation on thephenophase of walnut and varieties selection for late frostavoidance [J]. Deciduous Fruits, 2021, 53(1): 13-16.
[15] 王一峰, 赵淑玲, 王让军, 等. 甘肃成县核桃种质资源晚霜冻害调查[J]. 中国果树, 2020(1): 120-124.
[16] 仙米西努尔·克里木. 低温对新疆主要核桃品种枝条抗寒性生理指标的影响及抗冻措施[C]//中国环境科学学会2022年科学技术年会论文集(三). 中国环境科学学会, 2022:706-713.
[17] 张敏欢, 王建成, 杨红兰, 等. 新疆野核桃种质对低温胁迫的生理响应[J]. 应用生态学报, 2020, 31(8): 2558-2566.
ZHANG M H, WANG J C, YANG H L, et al .. Physiologicalresponse of Xinjiang wild walnut germplasm to low temperaturestress [J]. Chin. J. Appl. Ecol., 2020, 31(8): 2558-2566.
[18] 武彦霞, 史高川, 田鑫, 等. 5个核桃优系叶片抗寒性指标分析[J]. 果树资源学报, 2022, 3(6): 25-26,37.
WU Y X, SHI G C, TIAN X, et al .. Leaf cold resistanceindexes analysis of five walnuts excellent individuals [J]. J.Fruit Resour., 2022, 3(6): 25-26,37.
[19] 王一峰, 赵淑玲, 王瀚, 等. 不同核桃种质展叶期抗寒性的综合评价[J]. 经济林研究, 2019, 37(1): 50-60.
WANG Y F, ZHAO S L, WANG H, et al .. Comprehensiveevaluation on cold resistance of different walnut germplasms atleaf-expansion period [J]. Non-wood For. Res., 2019, 37(1):50-60.
[20] 吴硕, 贾彦丽, 智福军. 低温胁迫下核桃枝条抗寒性综合评价[J]. 林业与生态科学, 2020, 35(3): 314-319.
WU S, JIA Y L, ZHI F J. Comprehensive evaluation of coldresistance of walnut branches under low temperature stress [J].For. Ecol. Sci., 2020, 35(3): 314-319.
[21] 任俊杰, 赵爽, 苏彦苹, 等. 春季低温胁迫对核桃抗氧化酶指标的影响[J]. 西北农林科技大学学报(自然科学版), 2016,44(3): 75-81.
REN J J, ZHAO S, SU Y P, et al .. Effects of low temperaturestress in spring on antioxidant indexes of walnuts [J]. J.Northwest A&F Univ. (Nat. Sci.), 2016, 44(3): 75-81.
[22] 相昆, 徐颖, 王新亮, 等. 低温胁迫对核桃枝条活性氧代谢的影响[J]. 江西农业学报, 2014, 26(1): 35-37.
XIANG K, XU Y, WANG X L, et al .. Effects of lowtemperature stress on active oxygen metabolism in walnutbranches [J]. Acta Agric. Jiangxi, 2014, 26(1): 35-37.
[23] 田景花, 王红霞, 张志华, 等. 低温逆境对不同核桃品种抗氧化系统及超微结构的影响[J]. 应用生态学报, 2015, 26(5):1320-1326.
TIAN J H, WANG H X, ZHANG Z H, et al .. Effects of chillingstress on antioxidant system and ultrastructure of walnutcultivars [J]. Chin. J. Appl. Ecol., 2015, 26(5): 1320-1326.
[24] 韩立群, 马凯, 李亚利, 等. 低温胁迫下美国核桃和中国新疆核桃抗寒性综合评价[J]. 新疆农业科学, 2017, 54(10):1804-1812.
HAN L Q, MA K, LI Y L, et al .. Comprehensive evaluation ofcold resistance of American walnut and China Xinjiang walnutunder low temperature stress [J]. Xinjiang Agric. Sci., 2017, 54(10): 1804-1812.
[25] 刘荣, 刘清国, 范建新, 等. 果树抗寒性生理生化及分子机理研究进展[J]. 分子植物育种, 2017, 15(5): 2028-2034.
LIU R, LIU Q G, FAN J X, et al .. Research advances in thecold-resistance physiology biochemical and molecular mechanismin fruit crops [J]. Mol. Plant Breed., 2017, 15(5): 2028-2034.
[26] YANG G Y, ZHANG W H, LIU Z X, et al .. Both JrWRKY2 andJrWRKY7 of Juglans regia mediate responses to abioticstresses and abscisic acid through formation of homodimersand interaction [J]. Plant Biol., 2017, 19(2): 268-278.
[27] YANG G Y, ZHANG W H, SUN Y D, et al .. Two novel WRKYgenes from Juglans regia, JrWRKY6 and JrWRKY53, areinvolved in abscisic acid-dependent stress responses [J]. Biol.Plantarum, 2017, 61(4): 611-621.
[28] XU F, DENG G, CHENG S, et al .. Molecular cloning,characterization and expression of the phenylalanine ammonialyasegene from Juglans regia [J]. Molecules, 2012, 17(7):7810-7823.
[29] 刘凯, 李津津, 王红霞, 等. 核桃JrJAZ 基因家族鉴定与转录表达分析[J]. 河北农业大学学报, 2021, 44(1): 41-50.
LIU K, LI J J, WANG H X, et al .. Identification andtranscriptional expression analysis of walnut JrJAZ gene family [J].J. Hebei Agric. Univ., 2021, 44(1): 41-50.
[30] QUAN S W, NIU J X, ZHOU L, et al.. Genome-wide identification,classification, expression and duplication analysis of GRAS familygenes in Juglans regia L. [J/OL]. Sci. Rep., 2019, 9(1): 11643[2022-10-16]. https://doi.org/10.1038/s41598-019-48287-x.
[31] 刘凯, 赵星哲, 王红霞, 等. 核桃JrGRAS 基因家族鉴定与响应低温胁迫基因筛选[J]. 河北农业大学学报, 2021, 44(2):27-40.
LIU K, ZHAO X Z, WANG H X, et al .. Identification of walnutJrGRAS gene family and screening of genes in responce to lowtemperature stress [J]. J. Hebei Agric. Univ., 2021, 44(2):27-40.
[32] 赵云. 喷施GA3和ETH对核桃休眠芽萌发及抗寒力的影响[D]. 太原:山西农业大学, 2016.
ZHAO Y. Effects of GA3 and ETH on bud germinate and coldresistance for dormant buds of walnut [D]. Taiyuan: ShanxiAgricultural University, 2016.
[33] TANG K, ZHAO L, REN Y Y, et al .. The transcription factorICE1 functions in cold stress response by binding to thepromoters of CBF and COR genes [J]. J. Integrat. Plant Biol.,2020, 62(3): 258-263.
[34] ZUO Z F, KANG H G, PARK M Y, et al .. Overexpression ofICE1, a regulator of cold-induced transcriptome, confers coldtolerance to transgenic Zoysia japonica [J]. J. Plant Biol., 2019,62(2): 137-146.
[35] 李津津, 刘凯, 王芳, 等. 核桃响应低温胁迫转录因子ICE1基因的克隆与分析[J]. 林业与生态科学, 2021, 36(3):293-300.
LI J J, LIU K, WANG F, et al .. Cloning and analysis of coldstress transcription factor ICE1 gene in Juglans regia [J]. For.Ecol. Sci., 2021, 36(3): 293-300.
[36] 徐丽, 陈新, 张力思, 等. 核桃JrCOR 基因的克隆及表达分析[J]. 山东农业科学, 2018, 50(10): 13-18.
XU L, CHEN X, ZHANG L S, et al .. Cloning and expression ofJrCOR gene from walnut [J]. Shandong Agric. Sci., 2018, 50(10): 13-18.
[37] RUBIO S, NORIEGA X, PEREZ F J. Abscisic acid (ABA) andlow temperatures synergistically increase the expression ofCBF/DREB1 transcription factors and cold-hardiness ingrapevine dormant buds [J]. Ann. Bot., 2019, 123(4): 681-689.
[38] 徐丽, 陈新, 张力思, 等. 核桃JrCBF 基因的克隆与表达和单核苷酸多态性分析[J]. 植物遗传资源学报, 2014, 15(2):320-326.
XU L, CHEN X, ZHANG L S, et al .. Molecular cloning,expression, and single nucleotide polymorphism analysis oftranscription factor gene JrCBF in Juglans regia L. [J]. J. PlantGenet. Resour., 2014, 15(2): 320-326.
[39] 宋静武, 彭磊. 核桃CBF 基因在低温胁迫中表达[J]. 湖北林业科技, 2019, 48(2): 7-13.
SONG J W, PENG L. Investigating the expression of CBF genein Juglans regia under cold stress [J]. Hubei For. Sci. Technol.,2019, 48(2): 7-13.
[40] 徐丽, 陈新, 魏海蓉, 等. 核桃Y2SK2型脱水素基因JrDHN 的克隆、表达和单核苷酸多态性分析[J]. 园艺学报, 2014, 41(8): 1573-1582.
XU L, CHEN X, WEI H R, et al .. Molecular cloning,expression, and single nucleotide polymorphism analysis oftypical Y2SK2 dehydratin in Juglans [J]. Acta Hortic. Sin.,2014, 41(8): 1573-1582.
[41] YANG G Y, XU Z G, PENG S B, et al .. In plantacharacterization of a tau class glutathione S-transferase genefrom Juglans regia (JrGSTTau1) involved in chilling tolerance [J].Plant Cell Rep., 2016, 35(3): 681-692.
[42] 姬新颖, 冯启科, 刘志月, 等. 核桃JrPUBs(U-box)基因家族鉴定与响应冷温胁迫基因筛选[J]. 河北农业大学学报,2022, 45(6): 68-80.
JI X Y, FENG Q K, LIU Z Y, et al .. Identification of walnutJrPUBs (U-box) gene family and screening of genes inresponce to chilling stress [J]. J. Hebei Agric. Univ., 2022,45(6): 68-80.
(责任编辑:胡立霞)
基金项目:北京林业大学热点追踪项目(2022BLRD07);国家重点研发计划项目(2019YFD1002401)。
